S48.2: Multiple mechanisms of population regulation: Contributions of site dependence, crowding, and age structure
Nicholas L. Rodenhouse1, Thomas W. Sherry2, & Richard T. Holmes3
1Department of Biological Sciences, Wellesley College, Wellesley, MA 02181,USA, fax 781 283 3642, e-mail
NRodenhouse@Wellesley.edu; 2Department of Ecology, Evolution, & Organismal Biology, Tulane University, New Orleans, LA 70118-5698, USA; 3Department of Biological Sciences, Dartmouth College, Hanover, NH 03755-3576, USA Rodenhouse, N.L., Sherry, T.W. & Holmes, R.T. 1999. Multiple mechanisms of population regulation: Contributions of site dependence, crowding, and age structure. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2939-2952. Johannesburg: BirdLife South Africa.Pre-emptive use of sites (e.g. territories) that differ in suitability for reproduction or survival can lead to regulation of population size in passerine birds. This Site Dependent Regulation (SDR) has two major features: (1) environmentally-caused heterogeneity among sites in suitability for reproduction and/or survival, and (2) pre-emptive site occupancy, which is the tendency for individuals to use the best site available and to shift subsequently to sites of higher suitability as they become available. SDR, however, probably operates together with other mechanisms generating negative feedback to regulate population size. These other mechanisms include crowding effects and intrinsic differences among individuals, e.g. age and/or breeding experience, which influence reproduction or survival. Simulation modelling of a Black-throated Blue Warbler Dendroica caerulescens population revealed: (1) that the relative contributions of the mechanisms shift as population sizes grow from small to near saturation of suitable habitat, and (2) that intrinsic differences among individuals, e.g. those associated with age structure, may greatly influence the level of breeding productivity achieved for a given distribution of site suitabilities. We propose that the relative contribution of each mechanism varies greatly depending on the life history characteristics of species and environmental circumstances. These mechanisms can be distinguished by field experiments. The ability to detect and predict their operation would aid conservation efforts by identifying the factors and conditions that promote population growth.
INTRODUCTION
Understanding how the size of passerine bird populations is regulated involves more than the search for density dependence that has characterised such studies for more than 30 years. Indeed, knowing whether density dependence occurs in a population is of little value unless the specific mechanisms generating the regulatory feedbacks are also identified (Krebs 1991). Elucidating such mechanisms is essential because these processes connect measurable behaviours at the level of the individual with population-level phenomena, e.g. regulation of population size. It is from environmental conditions and resources acting at the level of the individual that population dynamics arise (Murdoch & Nisbet 1996), and manipulating environmental conditions and resources may often be the most efficacious way of managing avian populations (see Green & Hirons 1991). Hence, to understand or manage avian populations it is essential to know how the structure of the environment and the behaviour of individuals act together to generate negative feedback on demographic performance that in turn affects species’ abundances.
Several different kinds of mechanisms can generate the negative feedback necessary for regulation of local population size (reviewed by Sutherland 1996, Rodenhouse et al. 1997). These fall into two groups: those emphasising the primacy of interactions among individuals (e.g. crowding, territoriality/buffer effect) and those emphasising the structure of the environment (e.g. source-sink, site dependence). These groups of mechanisms can also be distinguished by the spatial scale over which they operate. For example, crowding mechanisms are generated primarily from local agonistic interactions among neighbours, whereas source-sink dynamics involve the movement of individuals among multiple sub-populations that may be widely dispersed. Studies of regulatory mechanisms within each of these groups has largely focused on interactions among individuals, very small (i.e. local) or very large (i.e. regional) spatial scales, and on single mechanisms. Seldom has the simultaneous operation of multiple mechanisms been determined or synthesised. Yet, it is likely that several mechanisms often operate simultaneously (den Boer & Reddingius 1996).
This paper has two objectives: (1) to highlight the potentially large role of environmental heterogeneity in population regulation of passerine birds, and (2) to exemplify via simulation modelling how multiple mechanisms might act together to regulate population size. Our method for achieving each of these goals focuses on the mechanism of site dependence. This mechanism stems from the ideas of Andrewartha & Birch (1954), Brown (1969), Fretwell and Lucas (1970), Lømnicki (1988), Pulliam & Danielson (1991) and others. Because site dependence, so named, is not yet widely recognised as a mechanism of population regulation, we begin with a brief description of that mechanism.
Site dependent regulation of population sizeRegulation of population size for site dependent species occurs when individuals pre-emptively use sites that differ in suitability for reproduction and survival (Rodenhouse et al. 1997). Species may be considered site dependent when their reproduction or survival depends on the characteristics of a site that they occupy exclusively or as a breeding pair. Sites for most passerine species would be the breeding or wintering territories they occupy. Regulation by site dependence can occur because, as population size grows, the average suitability of the sites occupied declines, reducing average reproductive performance or survival. This mechanism is not an alternative to density dependence, which represents regulatory negative feedback generated by a variety of mechanisms (Berryman 1991), one of which is site dependence. Site dependence has two major features: (1) environmentally-caused heterogeneity among sites in suitability for reproduction and/or survival, and (2) pre-emptive site occupancy, with the tendency for individuals to select initially or move subsequently to sites of higher suitability as they become available. Pre-emptive occupancy means that the best sites are occupied first, that site holders are not usually displaced, and that neighbours do not influence each other’s performance directly or indirectly (Pulliam & Danielson 1991).
Modelling demonstrates that site dependence can generate regulatory negative feedback in the absence of local crowding effects (Rodenhouse et al. 1997). Furthermore, the near ubiquity of the two main features of site dependence--heterogeneity among sites and the prevalence of dominance relationships within populations (e.g. most populations are structured by age, breeding status, experience on a site, etc.) -- strongly suggest that site dependence is a plausible mechanism of population regulation for a wide variety of passerine species (see Rodenhouse et al. 1997). Although supporting evidence for this mechanism exists (e.g. Andrén 1990, Dhondt et al. 1992, see Rodenhouse et al. 1997) no comprehensive tests of this mechanism have yet been carried out. Thus is it important to examine the generality and effectiveness of this mechanism, and to consider what its contribution might be relative to that of other mechanisms.
Relative contributions to regulation of three mechanismsSite dependence, crowding and intrinsic differences among individuals (e.g. those associated with age and/or experience) each potentially generate regulatory negative feedback at the local spatial scale. An examination of the contexts in which each of these mechanisms could contribute to regulation will aid in understanding their prevalence and potential roles.
Site dependence
For passerine birds, site dependence would most likely contribute to regulation of population size for species that obtain all resources on their territories, including both resident and migratory species that are territorial in either wintering and/or breeding areas. The reproductive success and survival of such species is expected to be directly related to the environmental conditions and resources found on their territories. Differences among territories in environmental conditions (e.g. weather) and requisite resources often result in significantly different reproductive success and survival (e.g. Ligon & Ligon 1988, Komdeur 1992, Braden et al. 1997). Many passerine species, however, do not seem to show such uniform dependence on the characteristics of particular sites. For example, non-territorial and colonial species often forage in a common space. However, one factor, such as food availability, is seldom the sole determinant of reproductive success or survival (Rodenhouse & Holmes 1992, Sherry & Holmes 1995). Species using food resources exploitatively might nest in sites that differ in vulnerability to predation. Any such site-related differences might potentially generate site dependent negative feedback.
Site dependence may be muted by a failure of pre-emptive site use. If individuals do not disperse widely and examine many sites, the closest sites, which are not necessarily the best sites, might be occupied first. Evidence from radio tracking and genetic studies counters this argument. Although dispersal distances are poorly known for most resident and migrant passerines, even residents seem to disperse very broadly relative to the diameter of breeding sites (Koenig et al. 1996). For some, if not most, migratory passerines natal dispersal occurs on regional spatial scales (e.g. Graves 1997). Furthermore, examination of a large number of sites of different suitability, or even accurate assessment of site suitability before the breeding season begins, is not essential to sustain pre-emptive site selection. Because individuals are most likely to show fidelity to sites of higher suitability and have higher survival on those sites, good sites will more frequently be occupied than poor sites. Thus individuals will most often be recruited to poor sites and may shift onto good sites when these become available. The effect on regulation of deviations from perfect pre-emption (i.e. habitats occupied sequentially from best to worst) is likely to be small (Pulliam & Danielson 1991).
Crowding
Crowding mechanisms have received much study, and these mechanisms can generate regulation (reviewed by Sinclair 1989, Newton 1992). Indeed most studies of 'density dependence' are implicitly or explicitly studies of crowding mechanisms. Negative feedback can be generated by these mechanisms because as numbers of individuals per unit area increases, direct (e.g. fighting) and indirect interactions among individuals (e.g. via enhanced nest predation) result in reduced reproduction or survival. However, the relationship between population density and individual performance is seldom linear, and significant negative feedback via crowding is usually thought to occur only when habitats are near or at saturation density (Murray 1994).
Where populations saturate suitable habitat, site dependence might be expected to have minimal regulatory influence because crowding effects might be strongest. Such crowding effects can significantly reduce reproduction (e.g. Arcese & Smith 1988) and survival (e.g. McCleery & Perrins 1985). However, it is usually very difficult to identify which habitats are suitable for most species (Petit & Petit 1996) because suitability is a continuum and many suitable sites may not be occupied in any particular year. Thus, identifying when all suitable sites are occupied, i.e., when a habitat is saturated, can be problematic (Marra & Holmes 1997). In some cases, however, there is no doubt of saturation, e.g. on islands or in isolated patches of suitable habitat. Available evidence suggests that even when the habitat is unquestionably saturated, as for the Seychelles Warbler on Cousin Island (Komdeur 1992), differences in site suitability can contribute to patterns of dispersal, reproductive success and survival of adults and juveniles. Each of these key demographic traits potentially plays a role in population regulation.
Intrinsic differences among individuals
Intrinsic differences among individuals include those associated with age/experience and phenotypic differences such as maternal effects (see Watson & Moss 1994). Such differences might contribute to population regulation over a broader range of population densities than crowding mechanisms because intrinsic differences can occur at all population densities. Intrinsic differences would contribute to regulation, for example, if the proportion of individuals attaining low reproductive success or survival increases as population size increases. This relationship would be expected, if the proportion of young inexperienced breeders increased with population size and many of them produced offspring at below replacement levels. Many studies show that age and social status can strongly influence reproductive success (Sæther 1990, Forslund & Pärt 1995); however, at least one study (i.e., Noordwijk & van Balen 1988) suggests that such differences among individual great tits Parus major may contribute little to reproductive success.
Few studies have examined the role that intrinsic differences among individuals alone may play in population regulation (but see Watson et al. 1994) because this requires parsing the effects of difference among individuals, interactions among individuals (i.e. crowding) and the inherent suitability of the sites occupied. Lømnicki (1988) explored multiple scenarios by which such intrinsic differences among individuals might contribute to population regulation. However, the scenarios he examined assume that available habitat is saturated. Hence the scenarios he explored confound limitation of population size via habitat saturation with the regulatory effects of intrinsic differences among individuals. Habitat saturation is not requisite for regulation by either intrinsic differences or site dependence; however, these two mechanisms may often operate together in generating regulatory negative feedback. For example, experienced breeders on a site of high suitability might have significantly greater reproductive success than inexperienced breeders on the same site (e.g. Goodburn 1991). Such differences among individuals would amplify regulatory feedback due to differences site suitability (Rodenhouse et al. 1997).
Distinguishing the regulatory roles of intrinsic differences among individuals, site dependence, and crowding effects under field conditions will be difficult and time consuming. Hence, it seems reasonable to begin exploring the relative roles of these distinct mechanisms by modelling their effects, using data from a well-studied population.
METHODS
Study population: Black-throated Blue Warbler
The premise of this paper is that regulation of passerine bird populations may occur via multiple mechanisms operating simultaneously. To assess the relative contributions to regulation of site dependence, intrinsic differences among individuals, and crowding mechanisms, we focus on a population that we have monitored and studied experimentally for more than 15 years -- that of the Black-throated Blue Warbler Dendroica caerulescens in the Hubbard Brook Experimental Forest, West Thornton, N.H., USA. This species is an open cup nesting passerine which breeds on exclusive territories in deciduous and mixed deciduous-coniferous forests in northeastern North America (Holmes 1994, Holmes et al. 1996 and references therein).
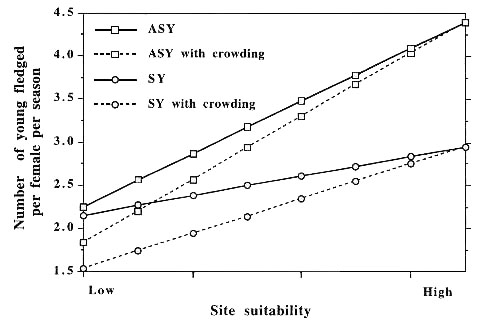
In BTBW, site dependent negative feedback on reproductive performance is proposed to arise as spatial differences in vegetation, food availability, weather, and predator abundances create differences in territory (i.e. site) suitability. We have demonstrated that each of these factors can significantly influence the breeding productivity of BTBW (Rodenhouse & Holmes 1992, Holmes et al. 1992, Holmes 1994), and we are currently testing the site dependence mechanism of population regulation on our study plots. Shrub density, in particular, is highly variable within northern hardwood forests, and it is associated with significant differences in the breeding performance of BTBW (Fig. 1).
Age-related intrinsic differences among individuals in breeding productivity and survival are found primarily between first time breeders (SY or yearling individuals) and experienced breeders (ASY or two years and older individuals). These intrinsic differences can be large; reproductive success of ASY individuals was almost twice that of SYs in the most suitable habitats (Holmes et al. 1996). Such intrinsic differences among individuals are common in passerines, particularly between first year and older breeders (reviewed by Sæther 1990, Forslund & Pärt 1995).
Crowding may also influence breeding performance in BTBWs. The density of breeding pairs can be up to three times as great in areas with high shrub density compared to where shrubs are sparse (Holmes et al. 1996). If crowding effects occur, we speculate that they will occur first for birds on the most suitable sites because these sites will be the first to fill when site selection is pre-emptive. However, it is likely that reductions in performance due to crowding will be greatest in the less suitable habitats (Fig. 1), because subordinate individuals with least breeding experience predominate on these sites. Most individuals occupying the best sites are experienced breeders--about 69% of individuals on plots with high shrub density are ASYs, but ASYs composed only 35% of individuals on low shrub density plots (Holmes et al. 1996). ASYs are presumed to be most able to defend their sites and nests as well as to care for their offspring. On the other hand, less experienced individuals are assumed to be affected more by direct or indirect effects associated with crowding.
Simulation modelThe model used to simulate the roles of these three regulatory mechanisms in a BTBW population is described in Rodenhouse et al. (1997). Shown schematically in Figure 2, the model includes eight different levels of site suitability that were determined by differences in reproductive performance and represented the range in breeding performance of BTBW found by Holmes et al. (1996). The model included 144 total breeding sites, and allowed the occurrence of floaters (i.e. non-breeding individuals). Sites were distributed unevenly among the eight levels of suitability, i.e., the model had fewer good than poor sites (Fig. 2). The distribution of sites among levels of suitability was chosen arbitrarily because we do not yet have field data on the distribution of site suitabilities for BTBW.
The model of Rodenhouse et al. (1997) was modified in two ways for the present study. Intrinsic differences among individuals and a crowding mechanism were added. Intrinsic differences among individuals were represented by age structure including ASY and SY individuals that differed in reproductive performance as shown in Figure 1. Parameter values were based on data from Holmes et al. (1996). Half of the initial 20 individuals were assigned to each age group for simulations including age structure. It was clear from studies on plots differing in shrub density (Holmes et al. 1996) that site suitability constrains the reproductive potential of both experienced (ASY) and inexperienced (SY) breeders. However, SYs benefited little from breeding on high shrub density plots (i.e. those of greatest suitability), whilst mean productivity of ASYs on these plots was about 25% greater than on the poorest ones. ASY pairs had about 75% greater productivity than SY pairs on the most suitable plots. These trends in the annual production of young were simulated in the model (see Fig. 1).
Crowding was represented by a direct feedback of total population size on annual production of young. The magnitude of this feedback differed among suitability levels (i.e. the difference between the solid and dashed lines representing ASYs or SYs in Fig. 1). Crowding reduced productivity only after each suitability level was half saturated (Fig. 3). As each suitability level filled beyond half saturation, production of young was progressively and linearly lowered so that it reached its lowest value at saturation density. It is important to note that once half saturation density was reached, productivity of all individuals in that suitability category was lowered as if all of these sites were located adjacent to one another. We chose this representation of crowding because it is the way that crowding is typically modelled in ideal free or despotic representations of habitat selection (Lømnicki 1988, Pulliam & Danielson 1991); however, this is not the only way that a crowding mechanism might operate. Analyses of long-term data comparing density to annual reproductive success suggest that a crowding mechanism may be operating for BTBW in the Hubbard Brook Experimental Forest (T.S. Sillett & R.T. Holmes, unpubl. data).
RESULTS
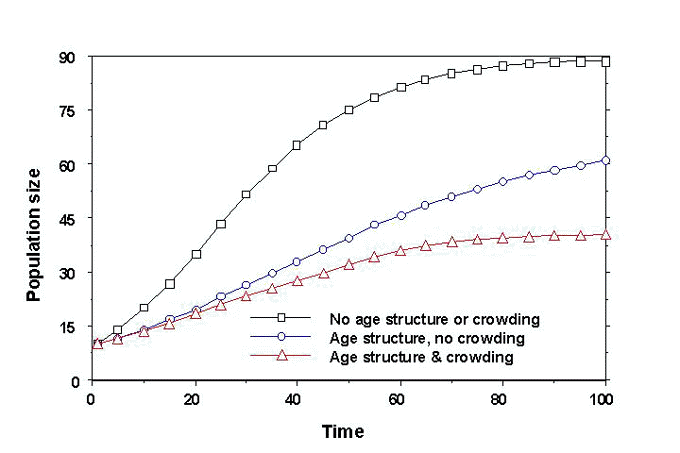
The baseline run of the model that included no intrinsic differences among individuals or crowding mechanism produced the most rapid population growth and highest equilibrium population size (Fig. 4). Rapid early growth occurred due to rapid filling of the most suitable sites and high productivity on those sites. Population growth slowed as poorer sites, including some with seasonal production of young below replacement level, were used by the expanding population. Adding intrinsic differences among individuals (i.e. age structure) to the model slowed growth in comparison to the baseline run and lowered equilibrium population size by about 20%. These reductions occurred because all individuals were forced to pass through the SY stage of low productivity. Adding a negative feedback due to crowding further slowed population growth and lowered equilibrium population size.
The crowding effect modelled was strong even at low population sizes because it reduced the suitability of the few best sites even when most sites were unoccupied. Whether a crowding mechanism such as this actually occurs for BTBWs is not yet clear. If one operates as it is modelled here, then it should have measurable effects over even relatively small differences in neighbourhood density. Note that crowding did not preclude pre-emptive selection of sites, i.e. sites were still filled from best to worst and site holders were not displaced. However, in contrast to site dependence operating as the sole mechanism, neighbours did affect each other’s productivity when the crowding mechanism was simulated.
Our simulations suggest two key points about regulation of passerine bird populations. First, that the relative contributions of different negative feedback mechanisms may shift as population sizes grow from small to near saturation of suitable habitat. For example, site dependence allowed rapid filling of the most suitable sites early in the simulations at a time when crowding and age structure forced slow population growth. Second, intrinsic differences among individuals, in this case associated with age structure, may strongly influence the level of breeding productivity achieved within a given distribution of site suitabilities. This implies that a young, relatively unproductive age class may severely limit population growth even when highly suitable sites are available. Discovering the causes of such low productivity in these early age classes may provide management options for species recovering from low population size.
DISCUSSIONThe simulations presented in this paper represent plausible scenarios for the contribution of multiple mechanisms to regulation of population size. If multiple regulatory mechanisms are operating, as exemplified in the third of these scenarios, then it is not surprising that passerine bird populations seem to be among the most tightly regulated (see Hanski & Tiainen 1989). However, each mechanism alone can be regulatory and the simulations provide few clues as to which mechanism or combination of mechanisms is likely operating under a given set of conditions in the field. Three scenarios aid in clarifying how the relative contributions of these mechanisms might differ depending upon environmental conditions and resources.
First, where many sites of high suitability and few poor ones occur, small populations would grow rapidly because of high reproductive success on the good sites (site dependence). As population size increases, the larger proportion of juvenile individuals would slow population growth (intrinsic differences among individuals). When high population density occurs, crowding effects might further slow growth or even cause declines in population size. Under this scenario, intrinsic differences among individuals and crowding effects are the primary regulatory forces with site dependence having minimal impact because of the high, largely uniform suitability of sites. However, even in this scenario that seems to minimise the potential impact of site dependence, stochastic demographic or environmental variation among years might create fluctuations in population size, and hence in the suitability-range of sites occupied, thereby enhancing the role of site dependence.
In the second scenario, a few good sites may be interspersed among many of lesser suitability. Under these conditions, site dependence may sustain population size at levels below which either intrinsic differences or crowding feedbacks occur. Intrinsic differences would have little impact because the proportion of, for example, juvenile individuals would remain small in a slowly growing population, and crowding would not occur unless individuals aggregated. Such aggregations, however, might be selected against if they resulted in reduced fitness of individuals in the aggregation as hypothesised for ideal despotic distributions (see Fretwell & Lucas 1970). This second scenario may be found among the numerous passerine species breeding within northern hardwoods forest that are sustained at relatively constant, low population densities, i.e., about 30 % of the migratory passerine species censused on the long-term study plots in the Hubbard Brook Experimental Forest (Holmes & Sherry 1988, and unpubl. data). This conjecture is based on the observations that few of these species breed in aggregations, and the distribution of annual breeding productivity for the most intensively studied species, the BTBW, is skewed. Among 225 BTBW pairs over 11 years, 26% produced no offspring and only 10% produced 6 or more (R.T. Holmes, unpubl. data). Such skewing is also reported for a variety of species by Newton (1989), and it would be expected if most sites are of intermediate or low suitability.
The third scenario represents species that breed in colonies or flocking species inhabiting semi-arid habitats that breed whenever and wherever conditions are suitable. Individuals of these species are typically non-territorial, forage exploitatively, and shift sites among breeding seasons. For such species, intrinsic differences among individuals may contribute the strongest negative feedback on population growth. Differences in foraging abilities, parenting skills and social dominance may contribute most to reproductive success. Because environmental favourability shifts greatly over time, population sizes, and hence the proportion of juveniles, may also change dramatically. Social dominance in particular may be important because it is associated with breeding position and reproductive success within colonies (e.g. Belestsky & Orians 1987, Shields et al. 1988, Picman & Isabelle 1995). Even in this scenario, however, other mechanisms may generate negative feedback on reproductive performance. Site dependence may play a role if sites on the edge of the colony have higher risk of predation than those in the centre, and crowding effects associated with food availability or parasitism may reduce reproductive success.
Each of these three scenarios may be plausible for some species, suggesting that none of the multiple mechanisms of regulation described can be ruled out a priori for most populations. Empirical data and our simulations lead us to expect that multiple sources of regulatory feedback will operate in general, and the relative contribution of each may vary greatly depending on the species’ life history characteristics and environmental circumstances. For example, species with weaker age/experience effects than BTBWs (e.g. those composed primarily of one age class) may be regulated predominantly by site dependence and crowding effects. Interestingly, these two mechanisms might have complementary roles in population regulation. Site dependence would allow most rapid recovery from small population size (because only the most suitable sites are occupied), and crowding effects could reduce growth when population size is large.
Noting that multiple mechanisms usually operate simultaneously does not mean that understanding regulation is hopelessly complex -- the different regulatory mechanisms identified in this paper can be distinguished and assessed. Regulation by site dependence is qualitatively different from that due to crowding mechanisms or intrinsic differences among individuals. These differences suggest experimental tests to distinguish the mechanisms, allowing assessment of their relative influence on population regulation. For example, site dependent negative feedback occurs as individuals pre-emptively use sites of differing suitability. Which particular individual occupies a particular site is not a part of this mechanism or of crowding mechanisms, but it is the essence of regulatory feedback generated by intrinsic differences among individuals. These differences among individuals result in some individuals achieving much higher productivity on certain sites than others. Tests distinguishing among these mechanisms could, therefore, focus on documenting site suitability, age of site occupant, site-specific breeding productivity and whether individuals influence directly or indirectly the fitness of their neighbours.
The focus of much previous research on population regulation has not been on mechanisms but rather on determining whether 'density dependence' (i.e. any negative feedback) occurs in bird populations (Bonsall et al. 1998). Evidence for 'density dependence' has been found or implied (reviewed by Sinclair 1989, Turchin 1995). However, the arguments and simulations presented in this paper suggest that understanding population regulation will require specific knowledge of the mechanisms by which such 'density dependence' occurs. Clarifying these mechanisms can provide information on the role of local interactions among individuals, of the spatial scale of regulation, and of the conditions under which regulation would be weakened or strengthened. Such knowledge can inform management practices for species conservation. For example, forest fragmentation is of great concern because it is associated with regional declines of some species. Such declines are due to fragmentation per se, loss of habitat or changes in habitat suitability. Fragmentation is considered harmful in each of these ways. But fragmentation could increase or decrease crowding, alter population structures and shift the distribution of site suitabilities. Such changes might eliminate regulation of population size or they could lead to even tighter regulation as the distribution of site suitabilities shifts (McPeek, M.A., Rodenhouse, N.L., Sherry, T.W. & Holmes, R.T., unpubl. MS.). Tighter regulation would reduce fluctuations in the size of small populations that might be particularly susceptible to chance events (Sæther et al. 1998). Experiments that distinguish among the mechanisms of regulation could turn these speculations into understanding and powerfully address conservation concerns for many passerine species.
ACKNOWLEDGEMENTSWe greatly appreciate the effort and input of the numerous field assistants and students who have worked with us at Hubbard Brook. This study was facilitated by the co-operation of the Northeast Forest Experiment Station, U. S. Forest Service, and particularly by C. W. Martin and C. Eagar. We thank J. N. M. Smith and S. Sillett for comments that helped us to improve this manuscript. This study was supported by National Science Foundation grants to N. L. Rodenhouse, T. W. Sherry, and R. T. Holmes.
REFERENCES
Andrén, H. 1990. Despotic distribution, unequal reproductive success and population regulation in the jay Garrulus glandarius L. Ecology 71:1796-1803.
Andrewartha, H.G. & Birch, L.C. 1954. The distribution and abundance of animals. Chicago; University of Chicago Press: 782pp.
Arcese, P. & Smith, J.N.M. 1988. Effects of population density and supplemental food on reproduction in song sparrows. Journal of Animal Ecology 57:119-136.
Beletsky, L.D. & Orians, G.H. 1989. Territoriality among male red-winged blackbirds. I. Site fidelity and movement patterns. Behavioral Ecology & Sociobiology 20: 21-34.
Berryman, A.A. 1991. Stabilization or regulation: what it all means! Oecologia 86: 140-143.
Bonsall, M.B., Jones, T.H. & Perry, J.N. 1998. Determinants of dynamics: population size, stability and persistence. Trends in Ecology & Evolution 13: 174-176.
den Boer, P.J. & Reddingius, J. 1996. Regulation and stabilization paradigms in population ecology. New York; Chapman & Hall: 397pp.
Braden, G.T., McKernan, R.L. & Powell, S.M. 1997. Association of within-territory vegetation characteristics and fitness components of California Gnatcatchers. Auk 114: 601-609.
Brown, J.L. 1969. Territorial behavior and population regulation in birds. Wilson Bulletin 81: 293-329.
Dhondt, A.A., Kempenaers, B. & Adriansen, F. 1992. Density dependent clutch size caused by habitat heterogeneity. Journal of Animal Ecology 61: 643-648.
Fretwell, S.D. & Lucas, H.L. Jr. 1970. On territorial behavior and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheoretica 19: 16-36.
Forslund, P. & Pärt, T. 1995. Age and reproduction in birds: hypotheses and tests. Trends in Ecology & Evolution 10: 374-378.
Goodburn, S.F. 1991. Territory quality or bird quality? Factors determining breeding success in the magpie Pica pica. Ibis 133: 85-90.
Graves, G.R. 1997. Geographic clines of age ratios of Black-throated Blue Warblers (Dendroica caerulescens). Ecology 78: 2524-2531.
Green, R.E. & Hirons, G.J.M. 1991. The relevance of population studies to the conservation of threatened birds. In: Perrins, C.M., Lebreton, J.-D. & Hirons, G.J.M. (eds) Bird population studies. Oxford; Oxford University Press: 594-633.
Hanski, I. & Tiainen, J. 1989. Bird ecology and Taylor’s variance-mean regression. Annales Zoologica Fennici 26: 213-217.
Holmes, R.T. 1994. Black-throated blue warbler (Dendroica caerulescens). In: Poole, A. & Gill, F. (eds) The birds of North America. Washington, D.C.; American Ornithologists' Union: 1-21.
Holmes, R.T. & Sherry, T.W. 1988. Assessing population trends of New Hampshire forest birds: local vs. regional patterns. Auk 105: 756-768.
Holmes, R.T., Sherry, T.W., Marra, P.P. & Petit, K.E. 1992. Multiple-brooding and annual productivity of a Neotropical migrant passerine, the Black-throated Blue Warbler (Dendroica caerulescens), in an unfragmented temperate forest. Auk 92: 321-333.
Holmes, R.T., Marra, P.P. & Sherry, T.W. 1996. Habitat-specific demography of black-throated blue warblers (Dendroica caerulescens): implications for population dynamics. Journal of Animal Ecology 65: 183-195.
Komdeur, J. 1992. Importance of habitat saturation and territory quality for evolution of cooperative breeding in the Seychelles warbler. Nature 358: 493-495.
Koenig, W.D., Van Vuren, D. & Hodge, P.N. 1996. Detectability, philopatry, and the distribution of dispersal distances in vertebrates. Trends in Ecology & Evolution 11: 514-517.
Krebs, C.J. 1991. The experimental paradigm and long-term population studies. Ibis 133 suppl. I: 3-8.
Ligon, J.D. & Ligon, S.H. 1988. Territory quality: key determinant of fitness in the group-living green woodhoopoe. In: Slobodchickoff, C. (ed) The ecology of social behavior. San Diego; Academic Press: 229-253.
Lømnicki, A. 1988. Population ecology of individuals. Princeton; Princeton University Press: 223pp.
Marra, P.P. & Holmes, R.T. 1997. Avian removal experiments: do they test for habitat saturation or female availability? Ecology 78: 947-952.
McCleery, R.H. & Perrins, C.M. 1985. Territory size, reproductive success and population dynamics in the great tit, Parus major. In: Silby, R.M. & Smith, R.H. (eds) Behavioral ecology. Oxford; Blackwell Scientific Publications: 353-373.
Murdoch, W.W. & Nisbet, R.M. 1996. Frontiers of population ecology. In: Floyd, R.B., Sheppard, A.W. & De Barro, P.J. (eds) Frontiers of population ecology. Collingwood; CSIRO Publishing: 27-43.
Murray, B.G. Jr. 1994. On density dependence. Oikos 69: 520-523.
Newton, I. 1989. Synthesis. In: Newton, I. (ed). Lifetime reproduction in birds. New York; Academic Press: 441-469.
Newton, I. 1992. Experiments on limitation of bird numbers by territorial behavior. Biological Reviews 67: 129-173.
Noordwijk, A.J. van & van Balen, J.H. 1988. The great tit, Parus major. In: Clutton-Brock, T.H. (ed). Reproductive success. Chicago; University of Chicago Press: 119-135.
Petit, L.J. & Petit, D.R. 1996. Factors governing habitat selection by Prothonotary Warblers: field tests of the Fretwell-Lucas models. Ecological Monographs 66: 367-387.
Picman, J. & Isabelle, A. 1995. Sources of nesting mortality and correlates of nesting success in Yellow-headed Blackbirds. Auk 112: 183-191.
Pulliam, H.R. & Danielson, B.J. 1991. Sources, sinks, and habitat selection: a landscape perspective on population dynamics. American Naturalist 137 (Supplement): S50-S66.
Rodenhouse, N.L. & Holmes, R.T. 1992. Results of experimental and natural food reductions for breeding Black-throated Blue Warblers. Ecology 73: 357-372.
Rodenhouse, N.L., Sherry, T.W. & Holmes, R.T. 1997. Site-dependent regulation of population size: a new synthesis. Ecology 78: 2025-2042.
Saether, B.-E. 1990. Age-specific variation in reproductive performance of birds. Current Ornithology 7: 252-283.
Sæther, B.-E., Engen, S., Islam, A., McCleery, R. & C. Perrins. 1998. Environmental stochasticity and extinction risk in a population of a small songbird, the Great Tit. American Naturalist 151: 441-450.
Sherry, T.W. & Holmes, R.T. 1995. Summer versus winter limitation of populations: what are the issues and what is the evidence? In: Martin, T.E. & Finch, D.M. (eds) Ecology and management of Neotropical migratory birds. New York; Oxford University Press: 85-120.
Shields, W.M., Crook, J.R., Hebblethwaite, M.L. & Wiles-Ehmann, S.S. 1988. Ideal free coloniality in the swallows. In: Slobodchikoff, C.N. (ed) The ecology of social behavior. San Diego; Academic Press: 189-228.
Sinclair, A.R.E. 1989. Population regulation in animals. Symposium of the British Ecological Society 28: 197-241.
Sutherland, W.J. 1996. From individual behaviour to population ecology. Oxford; Oxford University Press: 213pp.
Turchin, P. 1995. Population regulation: old arguments and a new synthesis. In: Cappuccino, N. & Price, P.W. (eds) Population dynamics: new approaches and synthesis. San Diego; Academic Press: 19-40.
Watson, A., Moss, R., Parr, R., Mountford, M.D. & Rothery, P. 1994. Kin landownership, differential aggression between kin and non-kin, and population fluctuations in the Red Grouse. Journal of Animal Ecology 63: 39-50.
Fig. 1. Estimated mean number of young fledged per female per season as a function of site (i.e., territory) suitability for ASY (After Second Year, i.e., experienced breeders) and SY (Second Year; those breeding for the first time) Black-throated Blue Warblers. Dashed lines indicate one possible scenario for effects of crowding on annual production of fledglings, and hence, on population regulation. Because sites are filled from best to worst, better sites are affected first, and crowding reduces production of fledglings by SYs more than by ASYs. Values for the annual production of young and the effects of crowding were estimated from the means and variances in the annual production of fledglings on sites of high and low shrub density presented in Holmes et al. (1996).
Fig. 2. Schematic of the simulation model used to assess the regulatory potential of site-dependence for Black-throated Blue Warblers. Eight levels of breeding site (i.e., territory) suitability are illustrated. Levels are sequentially occupied from most to least suitable as population size grows. Each suitability level includes a different number of breeding sites indicated by the number in each box (e.g. n = 12 for level 6). Sites of high suitability are rare relative to those of lesser suitability.
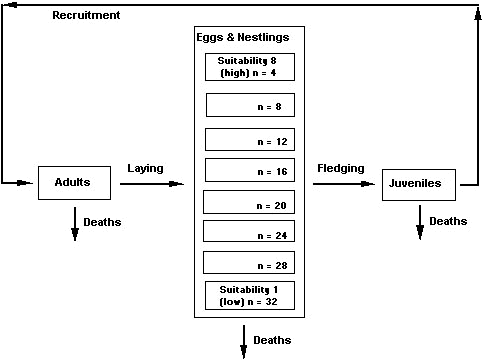
Fig. 3. Effects of crowding on
the number of young fledged per season simulated for ASY and SY pairs. Note that crowding
was assumed to lower annual production of young in a linear manner when more than half the
total number of sites in a suitability level were occupied (indicated by the arrow on the
x axis). We assumed also that crowding affected SYs more than ASYs (see text). This
pattern of reduction in annual production of young was simulated for each other level of
suitability (only level six with 12 sites is shown in this figure); however, the amount of
reduction differed among suitability levels. The amount of reduction simulated for each
level can be found in Fig. 1., where it is the difference between the solid and dashed
lines representing ASYs or SYs.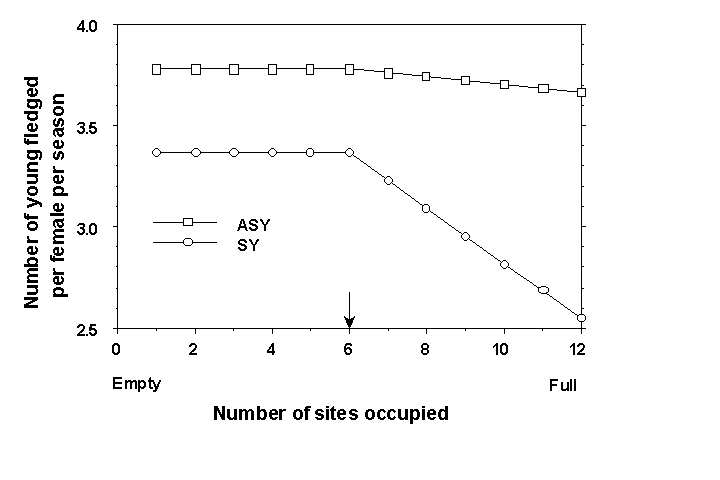
Fig. 4. Trajectories
of simulated Black-throated Blue Warbler populations affected by multiple regulatory
mechanisms: (o) site dependence with all individuals
alike but on sites of different suitability; (¡) site
dependence with age structure (ASY, SY differences as shown in Fig. 1), representing
intrinsic differences among individuals; and (Ñ ) both
of these mechanisms plus crowding as represented in Fig. 3. The simulation model used is
described in Rodenhouse et al. (1997).