S48.1: Behavioural mechanisms of population regulation in passerines
William J. Sutherland
School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, UK, fax 44 1603 592250, e-mail w.sutherland@uea.ac.uk
Sutherland, W.J. 1999. Behavioural mechanisms of population regulation in passerines. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2933-2938. Johannesburg: BirdLife South Africa.Density dependence is an essential process in population regulation and is important for understanding the consequences of habitat loss and habitat deterioration. The pattern of density dependence can be determined from models of behaviour. The relationship between mortality and density is expected to depend upon the rate of depletion and the nature of interference competition. The relationship between breeding output per female and density is likely to depend upon the variation in territory quality, the threshold territory quality at which individuals refrain from breeding, and the capacity to distinguish good from poor quality territories. The theory and evidence for these processes are reviewed.
INTRODUCTION
There has been considerable interest in linking behaviour and population ecology, but much of this work on birds has been on wading birds (e.g. Goss-Custard et al. 1995a,b) or geese (e.g. Percival et al. 1996, 1998). In this paper I suggest that there is considerable potential in using this approach for studying passerines. Many passerine species are good subjects for such an approach: their populations often have a high turnover so that it is easy to study births and deaths, passerines are often relatively easy to catch and mark individually, many nest in boxes making studies particularly easy (although introducing other artefacts), and the greatest advantage is that they are relatively easy to manipulate, for example, by adjusting the food supply or nest box density.
The global patterns of changes in the abundance of different habitats and habitat modification due, for example, to deforestation, agricultural changes, desertification, and climatic change obviously have affected almost all passerine species. Most species have experienced habitat loss and habitat deterioration although for some their area of suitable habitat will have increased or the habitat may have become more suitable for their requirements. Three well known conservation examples of changes in passerine populations are the declines due to logging and other forms of habitat loss on scarce tropical species (Collar et al. 1994), the decline in agricultural passerines resulting from more intensive agricultural practices (Fuller et al. 1995), and the much debated changes in populations of migrant passerines in North America (Peterjohn et al. 1995).
The importance of density dependence
Population size depends upon the interaction of density dependent and density independent processes (Williamson 1972). It thus follows that if these change then the population is likely to respond. From knowledge of density dependence it is possible to predict the decline in populations due to habitat loss (Sutherland 1996a) and habitat deterioration (Sutherland in press). The key component to this approach is that a loss of habitat or a decline in the quality of a patch of habitat will decrease the area of suitable habitat. If the increased density in the remaining area results in higher starvation, or if breeding habitat is lost, then the population will decline to a new equilibrium level. If there in no change in the birth or death rates for the same population size as a result of the population having to occupy a smaller area, then there will be no change in population. It is thus clearly important to be able to predict the relationships of birth and death rates to density, both as a key ecological process, but also for understanding the response to environmental change.
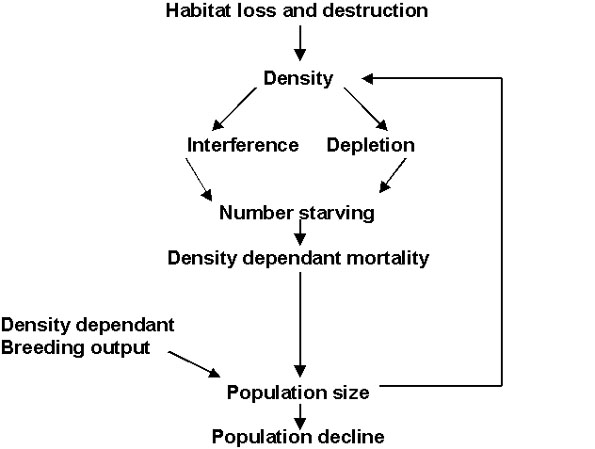
Density dependence can be studied by direct field studies (Newton 1998). An alternative approach is to use detailed studies of behaviour to determine the nature of the density dependent responses (Sutherland 1996b). One advantage of this latter approach is that it is possible to predict the response to novel conditions. As shown in Figure 1, the relationship between loss of winter habitat and population decline can be linked to the foraging behaviour. The density determines the importance of the interference and depletion which in turn determines the extent of density dependence. Similarly behavioural processes in the breeding grounds, and especially territorial behaviour, will determine the breeding output. In this paper, I review how the behavioural components may act in passerines. The three key components are interference, depletion and territoriality.
Interference
Interference is the reversible decline in intake rate resulting from the presence of others. Interference has been known for a long time from studies of parasitoids (Hassell & Varley 1969). Amongst vertebrates, most studies have been on wading birds especially Oystercatchers (e.g. Sutherland & Koene 1982). Amongst passerines, this phenomena has been shown for Snow Buntings Plectrophenix nivalis (Dolman 1995a) in which displacement over food patches determines the interference, and Blackbirds Turdus merula (Cresswell 1997) in which monitoring other individuals to avoid aggression reduces intake rate at high densities. Interference not only reduces local intake rate but is important in determining the distribution of individuals so that some individuals will then be in patches with lower prey density (Sutherland 1983).
DepletionThe number of individuals an area can hold can be related to the prey density and intake rate (Sutherland & Anderson 1993). A classic early example of the manner in which depletion can lead to density dependence is given by Gibb’s (1958, 1960) study of the Coal Tit Parus ater. Predation on the eucosmid moth Ernarmonia conicolana was higher in patches of higher density, and removed between 54 and 60% of the pupae. The survival of Coal Tits was positively correlated to the invertebrate stock over four years. The mixture of high depletion and a strong relationship between food supply and survival means that density dependence is very likely.
Dolman (1995b) reviewed all cases of depletion by vertebrates that he could find. This review suggests that passerines often deplete a large fraction of their prey. Silvereyes Zosterops lateralis deplete 40-67% of the codling moth cocoons Cydia pomonella (Wearing 1975), Starlings Sturnus vulgaris remove 42-57% of grass grubs Costelytra zealandica (East and Pottinger 1975) Great Tits Parus major take between 3 and 37% of various moth species (Tinbergen 1949), Goldcrests Regulus regulus take 23% of arboreal spiders (Askenmo et al 1977), Tree Sparrows Passer montanus take 11% of weed seeds (Wiens & Dyer 1977) and Pine Siskins Carduelis pinus take on average 58% of winter moth larvae Operophtera brumata (Roland et al 1986). In most cases it is clear that depletion is sufficiently high that there cannot be a considerable increase in population size. Furthermore, it is likely that in these cases there is variation in prey availability, for example with some seeds or invertebrates deeper in the soil, or that some individuals are better camouflaged so that subsequent prey will be increasingly harder to locate.
As an example of the consequences of a reduction in prey density, Rodenhouse & Holmes (1992) carried out an experimental reduction of caterpillar densities and showed that Black-throated Blue Warblers Dendroica caerulescens made fewer nesting attempts, although the clutch size, hatching success, and number of fledglings per nest were unaffected. Similarly, when comparing natural variation in caterpillar density between years, the strongest consequence of low food densities was a reduction in the number of breeding attempts.
Territorial behaviourThe occupancy of the better quality territories at low population density but a wider range of territory qualities at high density, as shown in great tits Parus major by Krebs (1971), will result in density dependent breeding output (Dhondt et al. 1992, Rodenhouse et al. 1997).
In many species of passerines some individuals do not breed even though capable of doing so and these have been referred to as 'floaters' (Smith & Arcese 1989). For example, in the Seychelles Magpie Robin Copsychus sechellarum on Fregate there are only about eleven territories. Even in a population of about twenty birds, some individuals are floaters. When these non-breeding birds were moved to an adjacent island (for conservation reasons) they then started to breed (Komdeur 1996). Newton (1998) tabulated removal studies of territorial passerines in the spring. Of the 34 studies, involving 23 species, in 21 one species was replaced while in 8 both sexes were replaced. Of these studies, in five it was known with certainty that some of these replacements came from floating individuals.
Floaters may occur even though there are empty areas of habitat that could be occupied. One explanation of this phenomenon is that individuals are refraining from occupying poor habitat in order to be free to occupy a better territory when one becomes available. Kokko & Sutherland (in press) model this process by considering a range of territories varying in quality, such that the best quality territory results in the highest reproductive success. It is assumed that the highest territories are occupied first and that individuals cannot swap territories once one has been selected.
The objective of this model is to calculate the minimum threshold territory quality at which an individual will decide not to breed but will instead wait for a better territory to become free. This is a game theory problem as the optimal decision depends upon the threshold decision of others, as this threshold determines the number of floaters who will be competing for territories.
The result of the game theory model is surprising: the threshold territory quality selected by individuals is the same as the threshold that maximises the number of floaters in the population. A low threshold means that all individuals breed so that the population is high but very few float. A high threshold means that the population is small, as only the best territories are occupied, these produce many young but as a result of the high threshold a high fraction float. Because population size is small, the absolute number of floaters is low. At an intermediate threshold the number of floaters is highest and it turns out that this threshold is exactly the optimal for individuals to gain the highest lifetime reproductive success.
This rule, that the threshold territory quality at which individuals refrain from breeding is the same as that which results in the highest number of floaters, is robust to a range of assumptions. Thus the same result applies whether the queue is strict, with the oldest occupant obtaining the next free territory, or lax, with any individual obtaining the next territory. Similarly it does not matter whether the queuing is for a given territory (as might be the case if helpers occupy the next occupancy) or for any territory, which would be the case if floaters moved through the area looking for vacant spaces.
This territoriality model can be used to examine the nature of density dependence (Kokko and Sutherland in press) by considering both how the breeding output from the different territories, and the number of individuals that decide not to breed, change with population size.
DISCUSSIONThere is great potential to measure some of the components of density dependence and apply the framework describe here. Interference is particularly straightforward to measure yet we have surprisingly few estimates of this important process. There are many of models of interference and it is clear that the key advance that is needed is an assessment of the main biological processes that determine interference and thus how they should be modelled. Depletion is also reasonably easy to quantify.
An important future area for research is the manner in which passerines sample areas. One of the key assumptions of the territoriality model is that individuals have perfect knowledge of habitat quality. The poorer the knowledge the weaker the density dependence will be (Pulliam & Danielson 1991). Territorial assessment will be a particular problem if quality is largely due to the risk of predation, which may be difficult for settling individuals to detect, or if settlement takes place before the main prey are present (Orians & Wittenberger 1991).
ACKNOWLEDGEMENTSI thank Richard Holmes and Jamie Smith for inviting me to talk at this meeting and for useful comments on an earlier draft.
REFERENCESAskenmo, C., Bromssen, von A., Ekman, J. & Janson, C. 1977. Impact of some wintering birds on spider abundance in spruce. Oikos 28: 90-94.
Collar, N.J., Crosby, M.J. & Stattersfield, A.J. 1994. Birds to Watch Vol 2: The World List of Threatened Birds. Birdlife Conservation Series. No 4. Birdlife International, Cambridge.
Cresswell, W. 1997. Interference competition at low competitor densities in blackbirds Turdus merula. Journal of Animal Ecology 66: 461-471.
Dhondt, A.A., Kempenaers, B & Adriansen, F. 1992. Density-dependent clutch size caused by habitat heterogeneity. Journal of Animal Ecology 61: 643-648.
Dolman, P. 1995a. The intensity of interference varies with resource density: evidence from a field study with snow buntings, Plectrophenix nivalis. Oecologia 102: 511-514.
Dolman, P. 1995b. Behaviour-based population models. Unpublished PhD thesis. University of East Anglia, Norwich, U.K.
East, R. & Pottinger, R.P. 1975. Starling (Sturnus vulgaris L.) predation on grass grub (Coslelytra zealandica (White), Melolonthinae) populations in Canterbury. New Zealand Journal of Agricultural Research 18: 417-452.
Fuller, R.J., Gregory, R.D., Gibbons, D.W., Marchant, J.H., Wilson, J.D., Baillie, S.R. & Carter, N. 1995. Population declines and range contractions among lowland farmland birds in Britain. Conservation Biology 9: 1425-1441.
Gibb, J.A. 1958. Predation by tits and squirrels on the eucosmid Ernarmonia conicolana (Heyl) Journal of Animal Ecology 27: 375-96.
Gibb, J.A. 1960. Populations of tits and goldcrests and their food supply in pine plantations. Ibis 102: 163-208.
Goss-Custard, J.D., Caldow, R.G., Clarke, R.T., Durell, S.E.A. le V dit & Sutherland, W.J. 1995a. Deriving population parameters from individual variations in foraging behaviour I. Empirical game theory distribution model of oystercatchers Haematopus ostralegus feeding on mussels Mytilus edulis. Journal of Animal Ecology 64: 265-276.
Goss-Custard, J.D., Clarke, R.T., Durell, S.E.A. Le V. dit, Caldow, R.W.G. & Ens, B.J. 1995b. Population consequences of winter habitat loss in a migratory shorebird. I. Estimating model parameters. Journal of Applied Ecology 32: 337-351.
Hassell, M. P. & Varley, G.H. 1969. New inductive population model for insect parasites and its bearing on biological control. Nature 223: 1133-1137.
Kokko, H & Sutherland, W.J. (in press). Optimal floating strategies: consequences for density dependence and habitat loss. American Naturalist.
Komdeur, J. 1996. Breeding of the Seychelles magpie robin Copsychus sechellarum and its implications for its conservation. Ibis 138: 485-98.
Krebs, J.R. 1971. Territory and breeding density in the Great Tit Parus major L. Ecology 52: 2-22.
Newton, I. 1998. Population Limitation in Birds. Academic Press, London.
Orians, G.H. & Wittenberger, J.F. 1991. Spatial and temporal scales in habitat selection. American Naturalist 137: S29-49.
Percival, S.M., Sutherland, W.J. & Evans, P.R. 1996. A spatial depletion model of the responses on grazing wildfowl to changes in the availability of intertidal vegetation during autumn and winter. Journal of Applied Ecology 33: 979- 993.
Percival, S.M., Sutherland, W.J. & Evans, P.R. 1998. Intertidal habitat loss and wildfowl numbers: application of a spatial depletion model. Journal of Applied Ecology 35: 57-63.
Peterjohn, B.G., Sauer, J.R. & Robbins, C.S. 1995. Population trends from North American Breeding Bird Survey. In Martin, T.E. and Finch, D.M. (eds) Ecology and management of neotropical migratory birds. Oxford University Press, New York: 3-39.
Pulliam, H.R. & Danielson, B.J. 1991. Sources, sinks and habitat selection: a landscape perspective on population dynamics. American Naturalist 142: 42-58.
Rodenhouse, N.L., Sherry, T.W., & Holmes, R.T. (1997). Site-dependent regulation of population size: a new synthesis. Ecology 78: 2025-2042.
Rodenhouse, N.L. & Holmes, R.T. 1992. Results of experimental and natural food reductions for breeding black-throated blue warblers. Ecology 73: 357-372.
Roland, J., Hannon, S.J. & Smith, M.A. 1986. Foraging patterns of pine siskins and its influence on winter moth survival in an apple orchard. Oecologia 69: 47-52.
Smith, J.N.M. & Arcese, P. 1989. How fit are floaters? Consequences of alternative behaviour in a non-migratory sparrow. American Naturalist 133: 830-845.
Sutherland, W.J. 1983. Aggregation and the ideal free distribution. Journal of Animal Ecology 52: 821-828.
Sutherland, W.J. 1996a. Predicting the consequences of habitat loss for migratory populations. Proceeding of the Royal Society 263: 1325-1327.
Sutherland, W.J. 1996b. From individual behaviour to population ecology. Oxford University Press.
Sutherland, W.J. (in press). The effect of change in habitat quality on populations of migratory species. Journal of Applied Ecology.
Sutherland, W.J. & Anderson, C.W. 1993. Predicting the distribution of individuals and the consequences of habitat loss: the role of prey depletion. Journal of Theoretical Biology, 160: 223-230.
Sutherland, W.J. & Koene, P. 1982. Field estimates of the strength of interference between oystercatchers. Oecologia 55: 108-109.
Tinbergen, L. 1949. Bosvogels en insekten. Nederlandsch Boschbouw Tijdschrift 21: 91-104.
Wearing, C.H. 1975. Integrated control of apple pests in New Zealand. 2. Field estimation of fifth instar larval and pupal mortalities of codling moth by tagging with cobalt-58. New Zealand Journal of Zoology 2: 135-149.
Wiens, J.A. & Dyer, M.I. 1977. Assessing the potential impact of granivorous birds in ecosystems. In Pinowski, J. & Kendeigh, S.C. (eds) Granivorous birds in ecosystems. Cambridge University Press, Cambridge: 205-266.
Williamson, M. 1972. The analysis of Biological Populations. Edward Arnold, London.
Fig. 1. The relationship between winter habitat loss and population change.