Bruce M. Beehler1 & Andrew L. Mack2
1Division of Environment and Natural Resources, Counterpart International, 1200 18th St, NW, Washington, DC 20036; and Division of Birds, National Museum of Natural History, Smithsonian Institution, Washington, DC 20560, United States of America, e-mail bbeehler@counterpart.org; 2Department of Conservation Biology, Conservation International, 2501 M St., NW, Washington, DC 20037, United States of America, e-mail a.mack@conservation.org
Beehler, B.M. & Mack, A.L. 1999. Constraints to characterising spatial heterogeneity in a lowland forest avifauna in New Guinea. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2569-2579. Johannesburg: BirdLife South Africa.
Between 1979 and 1998 we conducted 10 avian surveys at six forest sites in the Lakekamu Basin of Papua New Guinea. Two hundred and five bird species were recorded in lowland alluvial forest of the Basin: 170 breeding residents, 14 regular nonbreeding visitors, 10 local nonbreeding vagrants, and 11 migrants from Asia or Australia. The list includes 156 species of forest-dwelling birds, 82 of which are restricted to forest interior. No more than 171 species have been recorded from any single site. Surveys of less than two months’ duration produced significantly incomplete species lists; the recommended minimum effort for comprehensive site-based surveys in lowland rainforest of Papua New Guinea is four months total effort spread over several seasons. Results from individual surveys were skewed by a range of effects, some of which include observer bias, seasonal changes in avian detectability, and habitat-based effects. Having accounted for these sampling biases, we then present evidence for local patchiness of distribution within the Basin. Examples of locally patchy species include the Common Paradise Kingfisher Tanysiptera galatea and the Rusty Pitohui Pitohui ferrugineus, which are abundant at most sites but absent or rare at others. Floristic and structural heterogeneity at several scales in the Basin forests is undoubtedly a factor influencing within-Basin distribution of forest birds, and yet the complexity of the pattern of forest heterogeneity has hindered our delineating the full nature and extent to which birds track local forest patchiness. Future work should focus on this relationship.
INTRODUCTION
Ornithologists and conservationists need objective measures of the composition and size of a breeding bird community in a fixed survey period. Volumes have been produced on bird census methodology (Ralph & Scott 1981, Bibby et al. 1992; Ralph et al. 1995), and the diversity of techniques developed exemplifies the struggle to perfect dependable methodologies. Most avian census and survey work has been carried out in the temperate zone with well-known and species-poor avifaunas. Sampling populations of forest bird faunas in tropical lowland humid forests presents a range of especially vexing challenges. These include a poor understanding of seasonal changes in avian behaviour, their movement patterns (local, regional, seasonal, or aseasonal), and the difficulty of determining the resident status of species encountered rarely during a field effort (e.g., Robinson et al. 1990). These challenges are hindered by the structurally complex and poorly documented forest habitats that serve as home to the very rich tropical avifaunas of typical lowland rainforests (Beehler et al. 1995, Oatham & Beehler 1998).
The situation on the island of New Guinea offers challenges that are perhaps typical of the tropics. In this paper we review what we know of the avifauna of Papua New Guinea’s Lakekamu Basin, expand on our earlier assessment of the nature of the challenges to objective field survey in such environments (see Beehler at al. 1995), and, finally, assess the nature of local biotic heterogeneity.
METHODS
Assisted by an array of colleagues (see below and Acknowledgements), we conducted 10 field surveys in the Lakekamu Basin between 1979 and 1998 with the express purpose of sampling forest bird populations. Field surveys ranged in duration from 10 to 130 days. Some results from several of these expeditions have already been published (Beehler et al. 1994, 1995, Mack 1998), but here we have added four datasets that allow us to address the questions posed above.
Study area
The Lakekamu Basin (Fig. 1) of south-central Papua New Guinea (here termed ‘the Basin’, see Beehler et al. 1995) is one of the largest undisturbed tracts of lowland alluvial forest in southern PNG (see also Beehler 1993, Mack 1998). It supports only a small and scattered human population that currently produces only localised disturbance to the forest and its biota. The centre of the Basin was apparently a lake at some period in the past. This core 500-km2 area, when viewed from the air or on topographic maps, stands out as being a nearly level plain entirely surrounded by the sharp relief of border hills to the west and outliers of the central cordillera to the north, east, and south. The surface is slightly tilted, with the higher parts c. 100 m asl, the lowest c.40 asl. The distinct physical isolation of this tract of alluvial forest is a main reason it was selected for study. It was correctly assumed that the forest would not be commercially logged in the short term, making it attractive as a long-term research site. It still offers excellent potential as a conservation area.
Botanically, the lower part of this alluvial tract contains various types of swamp and swamp forest vegetation, whereas the upper section of the Basin is better drained and thus cloaked in continuous rainforest. We focused our avifaunal studies in this continuously-forested tract, as it appeared to us to represent the ‘ideal’ of Papuan lowland rainforest (Paijmans 1975) with minimum human disturbance. In fact, our recent field observations and those of visiting botanists (Reich 1998, Takeuchi & Kulang 1998) indicate considerable heterogeneity in forest structure, apparently related to movement of watercourses and other disturbances, differences in drainage and soil type, and human activities over the last century or more.
The Basin is in a moderate rainfall zone that receives 3 to 4 m of rain per annum, with the main wet season during the period from December to April (Beehler et al. 1995). Except during the strongest droughts, the forest interior is always very humid even in the annual dry season. Exceptional was a severe El Niño event that occurred from April to November 1997, causing widespread desiccation of the forest in conjunction with localised anthropogenic burning. We will comment on the possible impacts of this event based on a survey in May 1998.
Survey sites and survey effort
We surveyed birds during 10 field sessions at six forest field sites. Two sites (Nagore and Ivimka) were sampled three times. Two pairs of sites were situated fewer than 5 km apart on a single watercourse (the Korr and Ivimka sites and the Sii North and Sii South sites; Fig. 1a). Distance between nearest neighbour sites ranged from 5 to 10 km, and the two most distant sites were c. 20 km apart (Fig. 1a). All of the sites were superficially similar – situated in primary forest by a watercourse in alluvial outwash of clearwater drainages that dominate the Basin. Each site lies within what appears to be a single large continuous expanse of forest of c. 20 x 25 km. The only obvious environmental discontinuities that separate sites are rivers and streams, the largest of which is c. 30 m wide (the Oreba River, known locally as the ‘Biaru’). Thus our assumption was that we were sampling local communities within a single Basin avifauna where birds are presumably free to move between sites.
The study sites were as follows: 1. Kakoro camp (surveyed by Beehler & Ninga Kawa on 6 to 18 September 1979), a mix of lightly disturbed original alluvial forest, seasonally inundated swamp forest, and small regenerating garden patches at the base of steep low foothills of the central range c. 3 km NW of Kakoro air field (Beehler et al. 1995). 2. Sii River North (surveyed by Beehler & Stephen G. Pruett-Jones on 9 to 18 July 1982; = the ‘Si River Site’ of Beehler & Pruett-Jones 1983; Beehler et al. 1995), flat alluvial old growth forest on the SE banks of the Sii River. 3. Sii River South (surveyed by Beehler, Charles Burg, John Sengo, & Cosmas Makamet on 9 to 21 July 1994), a flat silt plain of grand alluvial forest near the confluence of the Oreba and Sii Rivers. 4. Nagore River camp (surveyed by Beehler & Burg on 16 to 20 April 1992; by Beehler, Sengo, Chris Filardi, & Kurt Merg on 3 July to 7 August 1992; and by Merg, John Cole, Beehler, Filardi, & Robert Bino on 1 April to 10 August 1993), a stony outwash plain of the rocky clearwater Nagore River supporting heterogeneous lowland alluvial forest. 5. Korr River (surveyed by Roger Safford, Leslie Smart, Fred Atwood, Makamet, and Sengo on 8 October 1994 to 7 January 1995), mid-Basin alluvial forest along the eastern course of the Avi Avi River and, of any sites, the most distant from any hilly lands. 6. Ivimka camp (surveyed by Safford, Smart, Atwood, Sengo, & Makamet on 15 February to 24 March 1995, by Jon Hornbuckle on 23 June to 3 July 1995 (these first two datasets are combined for this analysis), surveyed by Mack & Paul Igag on 15 October to 12 December 1996 [see Mack & Igag 1998], and by Beehler & Makamet on 18 to 27 May 1998, forest structurally much like that at Nagore and with bordering hills that lead into the central cordillera. Although at least one census included upland forest (Mack & Igag 1998), the upland data have been excluded from this analysis. Censuses at all camps were confined to the lowland forest.
Field Methods
At each field site we attempted a comprehensive assessment of the local bird community through a combination of systematic mist-netting, point censuses, and a daily tally of all ornithological sightings made by the field team. This combined dataset provided presence/absence information plus a general indicator of relative abundance (for details see Beehler et al. 1995, Mack & Igag 1998). At each site, fieldwork occurred within a single tract of forest c. 2.5 km in radius, where trails were cut to gain access to different parts of the forest tract. Because of the difficulty of standardising data collection among multiple, independent fieldworkers over several years, we generally avoid discussion of relative abundance (except in terms of extremes) and instead focus on presence/absence in our within- and between-site comparisons.
Our dataset consists of coded species lists of 10 site-based surveys, Kakoro (1 survey), Sii North (1), Sii South (1), Nagore (3), Korr (1), and Ivimka (3). This is further broken into mist-netting and census lists for each site. Mist nets were run at all six sites, and point counts were employed at Ivimka. Each point count consisted of a 10-minute sample at sites 200 m apart along parallel transects separated by 1000m. Point counts were undertaken in the morning between 0630 and 1000 h (Mack 1998).
Botanical censuses consisted of tree enumeration studies: two one-hectare plots at Nagore, a single one-hectare plot at Sii South, and two one-hectare plots at Ivimka. These included all woody stems larger than 10 cm dbh (for details see Beehler 1994, Oatham & Beehler 1998, Reich 1998).
Faunal analysis
Species lists for the 10 surveys were compared to delineate presence-absence and mist-netting results were used to quantify abundance of a subset of the species present in the different survey sessions. In addition, each species was classified by apparent residency status (breeding resident, nonbreeding visitor, local vagrant, austral migrant, or northern migrant) and habitat use (forest, forest-edge, nonforest, or any combination of these). Analyses were made using Notebook II database (Pro-Tem Software) and Systat 7.01 (SPSS).
RESULTS
The Avifauna
During the 10 field surveys, 205 bird species were recorded in the Lakekamu Basin lowland forest (Appendix). This list includes the following: 170 breeding residents, 14 regular nonbreeding visitors, 10 non-breeding vagrants, 8 Australian annual migrants, and 3 northern annual migrants. Ecologically, this list of 205 species includes 156 species of forest-dwelling birds (birds that inhabit forest interior but that also may use nonforest habitats), 103 species that use forest-edge, and 57 species that use edge or nonforest habitats. Whereas 82 species are strictly forest interior species, 19 species are confined to nonforest habitats in the Basin (Appendix).
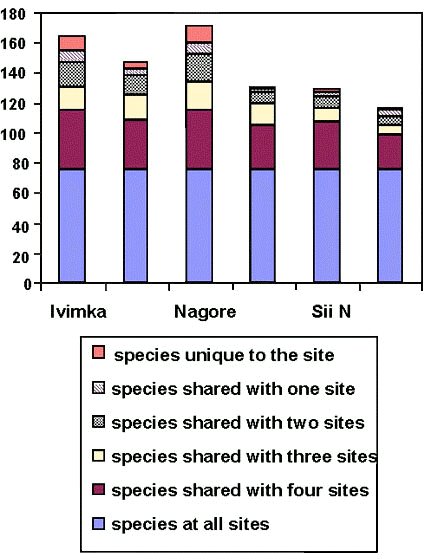
More than half (56%) of the species in the dataset were recorded on at least five of the six sites. Despite the presence of survey teams in the Basin for more than 400 days, there were still a large number of species (15%) that were recorded at only one site. The greater field efforts at Ivimka and Nagore resulted in over twice as many species rarely recorded (at just one or two sites) at those sites (Fig. 2).
Sampling effort
The species accumulation curve (or cumulative survey effort curve) shows our efforts were more than adequate to reveal most species that occur in the Basin (Fig. 3). For Ivimka, the point count effort curve levels off (Mack & Igag 1998), indicating this method had effectively sampled the diversity capable of being detected given the methods and observer skill level. In 330 minutes of counting, 932 individuals of 68 species were recorded (for more details see Mack & Igag 1998).
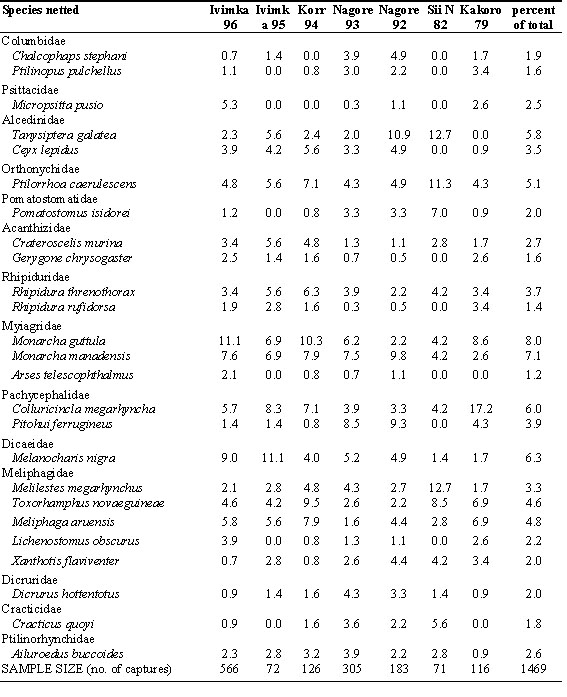
Excluding recaptures, 1469 birds representing 68 species were netted during seven netting sessions at five sites (Table 1). Because different levels of netting effort were employed, it is inadvisable to directly compare numbers netted, however comparisons can be made in terms of percentage of captures at a site. There is evidence of substantial differences in understory bird communities among the sites. Some species were abundant at some sites (e.g. Common Paradise-Kingfisher, Rusty Pitohui) and virtually absent at other sites (Table 1).
During 13 discrete census periods conducted at various times at the six sites, we recorded 108-171 species. The number of species recorded was positively correlated (rs=0.767, P<0.02) with census duration (Fig. 3).
Faunal similarity
We calculated turnover indices (Cody 1990) for the avian communities at each of the six sites, pooling all survey periods at those sites (Table 2). The index provides a unitless measure of similarity in all pairwise comparisons among the sites. Unity represents total dissimilarity (sharing no species) and zero would be complete similarity (sharing all species). Kakoro and Sii South were most dissimilar and Nagore and Ivimka most similar among the 15 pairwise comparisons. The differences in turnover indices were not correlated with distance between sites (rs= 0.384, P> 0.1). However, the turnover index was negatively correlated with observer effort among sites (rs= -0.746, P<0.01). The best-surveyed sites had the most similar species lists, indicating that the sites were part of a single Basin avifauna. Observer effort was defined as the sum of survey days at the two sites divided by the number of days at the best surveyed site (Nagore with 171 days). Thus effort scales between 2 (if both sites had been surveyed 171 days, to 0.012 if each site had only been sampled one day (see Table 2).
Forest Tree Heterogeneity
The flora of the Lakekamu Basin forest is biodiverse and heterogeneous at several spatial scales. At the largest scale, the lower and less-well-drained portions of the Basin (west and south-west) support swampy vegetation in 10 to 50 km2 patches quite distinct from the closed forests in the east where we conducted all of our work (see Fig. 1). We are currently unable to comment on the degree to which avian distributions track this large-scale heterogeneity because we have not visited these swamp patches.
At a more local scale, within the eastern closed forests of the Basin, there is evidence for inter-site heterogeneity from plot-based studies in the north (at Ivimka) and south (at Sii South and Nagore). The tree communities of the forests of Lakekamu Basin are species-rich, with one-hectare plots containing between 93 and 182 species of woody stems greater than 10 cm dbh (Oatham & Beehler 1998, Reich 1998). A most striking finding concerns the composition of the northern and southern plots. None of the 23 most abundant species on the southern plots ranked in the top ten in abundance for the two northern plots -- although they were only about 15 km apart. Furthermore, only 3 species of that top 23 for the southern plots were positively identified as occurring in the northern plots. Thus even in the well-drained upper part of the Basin there appears to be distinct communities in the southern (Nagore/Sii) and northern (Ivimka) sections. This is evidence for floristic patchiness at the scale of perhaps 1 to 10 km2.
At yet a smaller scale, it is notable that the two abutting one-hectare plots at Nagore (which thus shared a hundred-metre common boundary) were remarkably divergent as measured by presence or absence of tree species. The two plots supported 149 and 178 species, respectively, but the two plots combined included 252 species. Of this total, 103 tree species were confined to only one or the other of these two abutting plots. This is either evidence of local habitat patchiness at the c. one-hectare scale (presumably the product of plot history -- local wind-throws, stream movements, etc.) or that patch grain is larger than a hectare with high diversity within a patch.
Since, to date, we have surveyed no two plots that show even a moderate level of similarity in dominant species, it is clear that we have too few data to be able to characterise the nature of forest tree communities. We can say the forests of the Basin are a mosaic of species-rich communities in which tree species are typically distributed in very low densities (fewer than one tree per hectare). The communities are heterogeneous at at least three spatial scales, but the data are too few to objectively characterise this heterogeneity at a floristic level. Larger sample plots (> 4 ha.) may be required to reveal the nature of this arboreal heterogeneity.
The taxonomic heterogeneity of the forest is matched by its structural heterogeneity – largely a product of disturbance history. Within even small areas there are visible differences in bole size, frequency of buttresses, epiphyte loads, density of undergrowth, number of lianes, and gap dimensions (see Takeuchi & Kulang 1998). Delineation of such variation is beyond the scope of this paper, but there is sufficient evidence to conclude that the forest is structurally heterogeneous at a very local (1-5 ha) scale.
DISCUSSION
Intercontinental comparisons
The Lakekamu Basin breeding avifauna is impoverished compared to most continental tropical forest avifaunas. For example, ornithologists at the Cocha Cashu site in Manu National Park, Peru, have recorded an astounding 555 bird species (Robinson et al. 1990). Of these, 435 are regular residents (and wintering migrants) of the terrestrial habitats there (and thus exclude aerial, water-, and marsh-birds). In eastern Amazonia, near Manaus, Bierregaard (1990) recorded 352 species, with 260 of these forest-dwelling species. For M’Passa, Gabon, in a two square kilometre block, Brosset (1990) recorded 364 species, 175 of which are primary forest birds, 138 open habitat species, and 51 migrants. At Barro Colorado and adjacent Pipeline Road, Panama, Karr (1990) reports an overall bird list of 444 species -- 371 residents, 278 breeders, and 73 vagrants. Blake et al. (1990) report 237 species for the La Selva station in Costa Rica -- 178 forest breeding species and 59 forest nonbreeders. Making these comparisons objective is difficult because the variable scale and scope of the avifaunal assessments. Some are based on surveys on a single small forest plot, whereas others are based on large-scale multi-habitat analyses (Lakekamu tends to fall in the latter category because of the multiple plots and presence of riverine open habitats). Nonetheless, it is apparent that species diversity in the Basin and New Guinean lowland wet forest in general is lower than what is typical of similar forests in the Neotropics.
Efficacy of census method
Survey effort
The intensive efforts in the Basin have essentially revealed most of the species resident or regularly occurring there (Fig. 2). Three additional species were added in 1998 (Oriental Hobby Falco severus, Papuan Hawk-Owl Uroglaux dimorpha, and Yellow-gaped Meliphaga Meliphaga flavirictus). Given our knowledge of the ecology and geographic distribution of these three species, it is assumed these are breeding residents in the Basin, and hence each new survey effort continues to produce a very small number of additional resident breeding species. We believe that most of the remaining resident breeding species to be recorded in the Basin will be nonforest waterbirds such as rails and ardeids, although a few forest species remain expected. Doria’s Hawk Megatriorchis doriae, Papuan Hanging Parrot Loriculus aurantiifrons, and Barred Owlet-nightjar Aegotheles bennetti are possibilities, based on their geographic distributions. We expect considerable numbers of montane and hill forest wanderers to be added to the Basin avifauna as vagrants. Such vagrant species already on the list include the White-bibbed Fruit-Dove Ptilinopus rivoli, Scaly Thrush Zoothera dauma, and Mountain Drongo Chaetorhynchus papuensis.
Duration of our individual field surveys is positively correlated with the number of species recorded (Fig. 3). A one-month field session will produce a list of c. 130 species, mostly the commonest and most conspicuous species. However, longer field sessions (our maximum was 130 days) are required to approach a more complete list. Despite this, the total for the longest field session (171 spp.; Fig. 3) was still substantially lower than the pooled total for all sites (Fig. 2). This result agrees with that of Thiollay (this volume).
A substantial number of species were only recorded at one site (Fig. 4). A core group of 76 species was recorded at all sites and another 30-40 species were recorded at five of the six sites. The species recorded on a series of point counts at Ivimka nearly all came from this core group of widespread species.
Visual/audial daily tallies
Data from daily visual/audial tallies for 1998 give an indication of productivity of this sort of census method. Daily results ranged from 56 to 83 species, with individual totals ranging from 155 – 271 individuals counted. These are very useful, if rough, samples of the local bird community, based on compilation of diverse anecdotal data, compiled daily. Over a two to four month survey period, and when they include daily maximum counts, these datasets can provide useful local abundance measures.
Mist-netting
More variables constrain the netting results, including size and type of nets employed, the placement of the nets, and the number deployed. In the Lakekamu Basin, the typical net take is a bit less than 1 bird/day for a 12 metre 36 mm mesh net. For example, a 5-day effort with 20 nets at Nagore on 2-6 July 1993 captured a total of 78 individuals (including retraps) = 0.78 birds per 12 m net per day. This is about half that reported for Panama by Karr (1992). Number of species recorded by mist-netting in a single survey site (moving the nets regularly) over a field season typically is only about 50, compared to a total of 120 to 140 species for the visual/audial surveys. Netting did yield at least a few species not recorded otherwise on most of the censuses. However, as census duration increased, the number of species only recorded through netting decreased. Given the considerably greater investment required for netting, the return is marginal if one’s goal is rapid survey of community composition. Nonetheless, we note that netting is valuable for detecting rare, nonvocal species, and in situations where birdsong is not known by the survey team.
Constraints to sampling the avifauna
Seasonality
Because we have only one single-site dataset that extends from wet season to dry season (Nagore 1993), we have only limited ability to determine the degree of seasonal variation in species presence or detectability. We compare the four-week species list for 18 April - 16 May 1993 against the list for 12 July - 7 August 1993. The two lists differed considerably, perhaps because the broader census efforts that took place in the July-August segment. No obvious biological difference in species presence or detectability was noted except for the northern and southern migrants – whose movements are already well documented. Survey staffing and effort must be standardised and fine-tuned in order to provide data of the precision suitable for subtle seasonal changes in avian detectability or actual movement in and out of the site.
Year-to-year variation
Virtually the same survey crew worked at Nagore during the July-August seasons in 1992 and 1993. Comparing these two datasets permits us to pinpoint any evidence of inter-year variation in community composition. There was virtually none that could be attributable to anything other than obvious observer error (cryptic and poorly known species that became better known over time).
Supra-seasonal phenomena
In 1997 a severe El Niño event struck Papua New Guinea, causing widespread drought and famine. The rains failed for 7 months (April to October 1997) in the Lakekamu Basin. Fires burned forest in many places adjacent to human habitation. Virtually all of these were set by humans, intentionally or inadvertently. By the time of our most recent census in May 1998, the rains had arrived and the forest was again typically perhumid, and yet that survey produced odd anomalies that may have been the result of the long drought. Comparing the results of the earlier Ivimka surveys with that of the May 1998 survey, we find that the following three commonplace species recorded in 1995 and 1996 were not recorded in 1998: Southern Crowned Pigeon, Common Paradise-Kingfisher, and Blue-breasted Pitta Pitta erythrogaster. These three are species recorded throughout the Basin, typically easy to locate, and usually vocal. It is notable that all three obtain their food exclusively from the ground (although by differing methods); we cannot determine why the drought might have affected these species and not others, including other ground-foraging species.
Inter-site Avifauna Heterogeneity in the Basin
Turnover indices for the different sites within the Basin (Table 2) are fairly homogeneous. There are not any sites that stand out as radically different from the others. Similarity between sites was not correlated with distance between sites-- the closer sites were not more similar than sites further apart. However, there is a significant negative correlation between the turnover index and survey effort. The longer that sites were sampled the more similar their respective species lists became.
Examples of local patchiness
The two Sii River sites are a mere 3 km apart, on the same small river in continuous forest, but their ecological character differs. Sii North is close to low but rugged hills, and the river itself is rocky and shallow. Sii South is flat, with silty soils, and the river is slower and sandy-bottomed. The two exhibited slight but distinct differences in their avifauna lists of 129 and 131 species. Twenty-five species were unique to Sii North and 27 to Sii South. For the fruit doves (Ptilinopus) Sii North included two hill forest and montane species (White-bibbed Fruit-Dove P. rivoli and Coroneted Fruit-Dove P. coronulatus). By contrast, Sii South included two patchily distributed lowland forms absent from Sii North (Dwarf Fruit-Dove P. nanus and Orange-bellied Fruit-Dove P. iozonus). Additional hill forest forms in the Sii North plot include the Stout-billed Cuckoo-shrike Coracina caeruleogrisea and two upland lorikeets (Little Red Lorikeet Charmosyna pulchella and Pygmy Lorikeet C. wilhelminae), as well as a hill forest cuculid (White-crowned Koel Caliechthrus leucolophus). The Sii South site included three large eagles (New Guinea Harpy-Eagle Harpyopsis novaeguineae, Gurney’s Eagle Aquila gurneyi, and White-bellied Sea-Eagle Haliaeetus leucogaster), whereas Sii North included none. Finally, and somewhat mysteriously, the Sii South site did not include Meyer’s Friarbird Philemon meyeri, a common lowland forest species expected at all Lakekamu sites. Most other differences can be attributed to sampling artefacts; both sites were only censused for short periods of 10 and 13 days.
The Korr and Ivimka sites are situated along the same river (the east branch of the Avi Avi) c. 10 km apart. The ecological differences between the sites almost exactly match those of Sii North (like Ivimka) and Sii South (like Korr). Species lists for the two total 147 and 140, and ‘single-site’ species totalled 25 (Korr) and 18 (Ivimka 95). Avifaunally, the differences are similar to the Sii example, as well. There is a suite of upland species that appear on the Ivimka list but are absent from the Korr site, which is isolated from any hill forests (White-bibbed Fruit-Dove, Goldie’s Lorikeet Trichoglossus goldiei, Lilac-collared Parrot Geoffroyus simplex, Wallace’s Wren Sipodotus wallacei, River Flycatcher Monachella muelleriana, and Mountain Drongo). By contrast, the Korr site includes four water birds absent from the Ivimka list (Little Pied Cormorant Phalacrocorax melanoleuca, Great Egret Egretta alba, Red-necked Rail Rallina tricolor, and Bush-hen Amaurornis olivaceus).
The major avifaunal comparison of local communities must be between Nagore and Ivimka, both situated near hill forest and both bisected by rocky and swift-running rivers. We compare compiled lists for the two sites. Nagore included 171 species, 26 not found at Ivimka. Ivimka included 164 species, 19 not included at Nagore. Both of the lists of species found only at one site include small components of the hill forest fauna – Nagore with 3 unique hill species, Ivimka with 9 unique hill species. Otherwise, Nagore, with more open riverine habitat, supported more water and open country bird species (14 species unique to Nagore). By contrast, Ivimka uniquely supported only 3 open country and water bird species. These faunal differences are neither unexpected nor notable – they are related to proximity to either hill forest or open country habitats. Of significance is that the species lists from these two well-investigated sites were very similar, overall.
Most interesting of all, but also very difficult to document objectively, is the selective occurrence of key forest-dwelling species in the Basin. This can be termed 'local patchiness' [at a scale much smaller than that described by Diamond (1980)] and is a phenomenon that we can only begin to document here, but that should be addressed by additional focused fieldwork in the future. Some of it may be habitat selection within the mosaic of closed forest types that we are, as yet, unable to delineate. Some may be the product of remnant or small and decaying local populations. Needless to say, it is of significance to conservation planners, because certain forest-dwelling bird species may not be continuously distributed through large expanses of continuous humid forest.
To illustrate our point we begin with some counter-examples, the birds of paradise. Six species inhabit the Basin and all six have been recorded at all six survey sites (Appendix). Clearly the birds of paradise show no evidence of local patchiness. By contrast, the following 6 examples are notable as lowland forest bird species that unambiguously occur in some Basin forests and not others.
The Collared Imperial Pigeon Ducula muelleri is a poorly-known forest-dweller that appears to prefer riverine habitats (Coates 1985). It has been recorded only at the Korr site -- which, perhaps not coincidentally, is closest to the larger sections of the Lakekamu River system (Fig. 1). The Blue-black Kingfisher Halcyon nigrocyanea is a little-known denizen of forest-interior, with a reputation for preferring swampy forest (Coates 1985). In the Basin it is known from a single netted individual at Ivimka (Mack 1998). All of the Basin’s kingfishers are netted easily, so this species is evidently very rare and very localised. Perhaps it will be found to be commonplace in the western flooded portion of the Basin. The Little Kingfisher Alcedo pusilla has been recorded at only three of the six sites (Kakoro, Nagore, Korr). Its distribution is inexplicable. Likewise, the Hooded Pitta Pitta sordida, unlike its more abundant cousin the Blue-breasted Pitta, is known only from three sites (Kakoro, Sii North, and Korr). The Yellow-breasted Boatbill Macheirirhynchus flaviventer has been recorded at three of the six sites (Kakoro, Korr, Ivimka). It is typically a common flock-follower that is vocal and easy to detect. Finally, the strikingly vocal Black-sided Robin Poecilodryas hypoleuca was encountered only at Kakoro, Ivimka, and Korr. This bird’s voice rings through the forest at sites where it is found (it was widespread at Ivimka in 1998 and encountered dozens of times).
What characteristics are shared by this group of six patchy species? All inhabit forest interior (two also can be found at forest edge). All but the pigeon subsist on arthropods or small vertebrate prey. Three take their prey from the ground or from small watercourses (kingfishers and pitta). More study of this phenomenon is in order.
Final thoughts
Proving Residence
We estimate that as many as 30 or 40 hill and montane vagrants may be added to the Basin list over time, mainly because of the physical proximity of upland habitats to the this tract of lowland forest. Montane forest is but a few kilometres, as the crow flies, from the verge of the Basin (Fig. 1). This group of vagrant species is an additional hindrance to objective analysis of the Basin breeding avifauna. Unless one documents local breeding, how does one objectively determine residency? The same might be said for the Blue-black Kingfisher or Collared Imperial Pigeon. Are they residents? For rarely encountered species this will be difficult indeed.
Year-round censuses
Forest bird censuses in New Guinea should be repeated at intervals throughout the year. This is potentially possible for staff managing field stations like those at Ivimka or Crater Mountain in Papua New Guinea, but reliable censuses depend on presence of an observer knowledgeable of bird song. Replication of year-round censuses will allow us to address issues of seasonality of detectability vs. seasonality of presence – a critical issue to this sort of survey effort. Mist-netting censuses sample a relatively small subset of the resident avifauna, but can be undertaken by people unfamiliar with songs. Mist-netting can be standardised to determine population changes of easily-netted species (e.g., Blue-faced Parrot Finch Erythrura trichroa see Mack & Wright 1996) and provide additional useful data on avian moult or breeding cycles.
Knowledge of bird song
Without question, effective comprehensive survey of forest birds in lowland forests of New Guinea requires extensive knowledge of birds' songs and the calls. This is by no means a simple enterprise for, to date, there is no comprehensive sound library compiled for the region. Given the complexity and variability of the vocal repertoire of many species and the large areas where no recordings have been made, a single set of recordings for the country is not yet feasible. Instead, local or regional compilations will be necessary, and each regionally-based project should make concerted efforts to tape record and catalogue all bird sounds from the site. Those vocalisations that are unidentified should be noted and recorded, for later identification. Both of the authors freely admit that their knowledge of bird song for the Lakekamu Basin is incomplete. We suggest that sound-based bird identification in New Guinea, initially highlighted by Diamond (1972), remains a priority for creating a reliable basis for any future ornithological field assessments. The legacy of Ted Parker (Parker 1991) is not forgotten.
ACKNOWLEDGEMENTS
The governments of Papua New Guinea and Gulf and Central Provinces very kindly permitted the authors to conduct rainforest research in the Lakekamu Basin over the years 1979 through 1998. We thank the Research and Exploration Committee of the National Geographic Society, the Wildlife Conservation Society, and Conservation International for supporting our research in the Lakekamu Basin. In addition, the research benefited from grants from the MacArthur Foundation, the Biodiversity Conservation Network, the U.S. Agency for International Development, the Conservation, Food and Health Foundation, and the Papua New Guinea Biological Foundation. The hospitality of the four ethnic groups in the region, the Kurija-Kunimaipa, the Kamea, the Biaru, and Kovio peoples made possible our research on their traditional lands. Thanks also go to the following who assisted in the field and in home offices in Port Moresby and back in the United States: Carol Beehler, Yati Bun, Chuck Burg, Joseph Dumoi, Christopher Filardi, Julie Hagelin, Matai Kai, Aubita Kairo, Korau Kaiu, Cosmas Makamet, Kurt Merg, Mary Pearl, John Poia, Stephen G. Pruett-Jones, Harry Sakulas, John Sengo, Joseph Wiakabu, and Debra Wright.
REFERENCES
Beehler, B.M. (ed.) 1993. A biodiversity analysis for Papua New Guinea. Conservation needs assessment. Vol 2; Washington, D.C.; Biodiversity Support Program.
Beehler, B.M. 1994. Using village naturalists for treeplot biodiversity studies. Tropical Biodiversity 2: 333-338.
Beehler, B.M., Burg, C.G., Filardi, C. & Merg, K. 1994. Birds of the Lakekamu-Kunimaipa Basin. Muruk 3: 1-8.
Beehler, B.M. & Pruett-Jones, S.G. 1983. Display dispersion and diet of birds of paradise: a comparison of nine species. Behav. Ecol. and Sociobiol. 13: 229-238.
Beehler, B.M., Sengo, Filardi, C. & Merg, K. 1995. Documenting the lowland rainforest avifauna in Papua New Guinea -- effects of patchy distributions, survey effort, and methodology. Emu 95: 149-161.
Bibby, C.J., Burgess, N.D. & Hull, D.A. 1992. Bird census techniques. New York; Academic Press: 257pp.
Bierregaard, R.O. 1990. Species composition and trophic organization of the understory bird community in a Central American terra firme forest. In: Gentry, A.H. (ed.) Four neotropical rainforests. New Haven; Yale University Press: 217-236.
Blake, J.G., Stiles, F.G. & Loiselle, B.A. 1990. Birds of La Selva Biological Station: habitat use, trophic composition, and migrants. In: Gentry, A.H. (ed.) Four neotropical rainforests. New Haven; Yale University Press: 161-182.
Brosset, A. 1990. A long term study of the rain forest birds in M’Passa (Gabon). In: Keast, A.H. (ed.) Biogeography and ecology of forest bird communities. The Hague; SPB Academic Publishing: 259-274.
Coates, B. 1985. Birds of Papua New Guinea. Vol 1; Alderley, Queensland; Dove Publications: 464pp.
Cody, M.L. 1993. Bird diversity components within and between habitats in Australia. In Ricklefs, R. E. & Schluter, D. (eds) Species diversity in ecological communities. University of Chicago Press, Chicago: 147-158.
Diamond, J.M. 1972. Avifauna of the eastern highlands of New Guinea. Cambridge, Massachusetts; Nuttall Ornithological Club: 438pp.
Diamond, J.M. 1980. Why are so many tropical bird species distributed patchily with respect to available habitat? In Nöhring, R. (ed.) Proceedings of the XVII International Ornithological Congress. Berlin: 968-973.
Karr, J.R. 1990. The avifauna of Barro Colorado Island and the Pipeline Road, Panama. In: Gentry, A.H. (ed.) Four neotropical rainforests. New Haven; Yale University Press: 183-198.
Mack, A.L. & Wright, D.W. 1996 Birds of the Crater Mountain Biological Research Station, Papua New Guinea. Emu 96: 89-101.
Mack, A.L. (ed.) 1998. A biological assessment of the Lakekamu Basin, Papua New Guinea. RAP Working Paper No. 9. Washington, DC; Conservation International: 186pp.
Mack, A.L. & Igag, P. 1998. Birds. In: Mack, A. (ed.) A biological assessment of the Lakekamu Basin, Papua New Guinea. RAP Working Paper No. 9. Washington, D.C.; Conservation International: 62-67.
Oatham, M. & Beehler, B.M. 1998. Richness, taxonomic composition, and species patchiness in three lowland forest plots in Papua New Guinea. In: Dallmeier, F. & Comisky, J. (eds) Forest biodiversity research, monitoring and modeling: conceptual background and old world case studies. Casterton, U.K.: Parthenon Publishing: 649-668.
Paijmans, K. 1975. New Guinea vegetation. Amsterdam; Elsevier Scientific Publishing: 213pp.
Parker, T.A. III. 1991. On the use of tape recorders in avifaunal surveys. Auk 108: 443-444.
Ralph, C.J., Sauer, J.R. & Droege, S. (eds) 1995. Monitoring populations by point counts. Albany, California; U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: General Technical Report PSW-GTR-149: 187pp.
Ralph, C.J. & Scott, J.M. (eds) 1981. Estimating numbers of terrestrial birds. Studies in Avian Biology 6: 630pp.
Reich, J.A. 1998. Vegetation, Part 1: a comparison of two one-hectare tree plots in the Lakekamu Basin. In: Mack, A. (ed.) A biological assessment of the Lakekamu Basin, Papua New Guinea. RAP Working Paper No. 9. Washington, D.C.; Conservation International: 23-33.
Robinson, S.K., Terborgh, J. & Munn, C.A. 1990. Lowland tropical forest bird communities of a site in western Amazonia. In: Keast, A.H. (ed.) Biogeography and ecology of forest bird communities. The Hague; SPB Academic Publishing: 229-258.
Takeuchi, W. & Kulang, J. 1998. Vegetation, Part II: Botanical Survey. In: Mack, A.L. (ed.) 1998. A biological assessment of the Lakekamu Basin, Papua New Guinea. RAP Working Paper No. 9. Conservation International, Washington, DC; Conservation International: 34-37.
Table 1. The twenty-five most commonly-netted bird species in the Lakekamu Basin. Captures are expressed as percent (%) of the captures (excluding recaptures) during each of seven netting sessions at five site in different years (1979-1996). Less frequently captured species are not listed here but were used in calculation of percentages at each site.
Table 2. Matrix of turnover indices (see Cody 1993 and results) below the diagonal and a measure of sampling effort above the diagonal (see results).
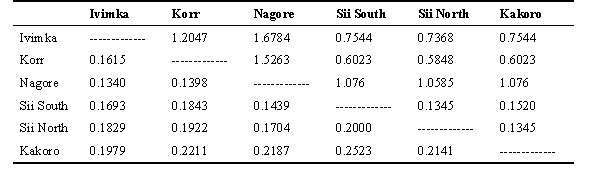
Fig. 1. (a) Lakekamu Basin, Papua New Guinea, showing survey sites (names displayed in all caps). (b) Location of the Basin within Papua New Guinea.
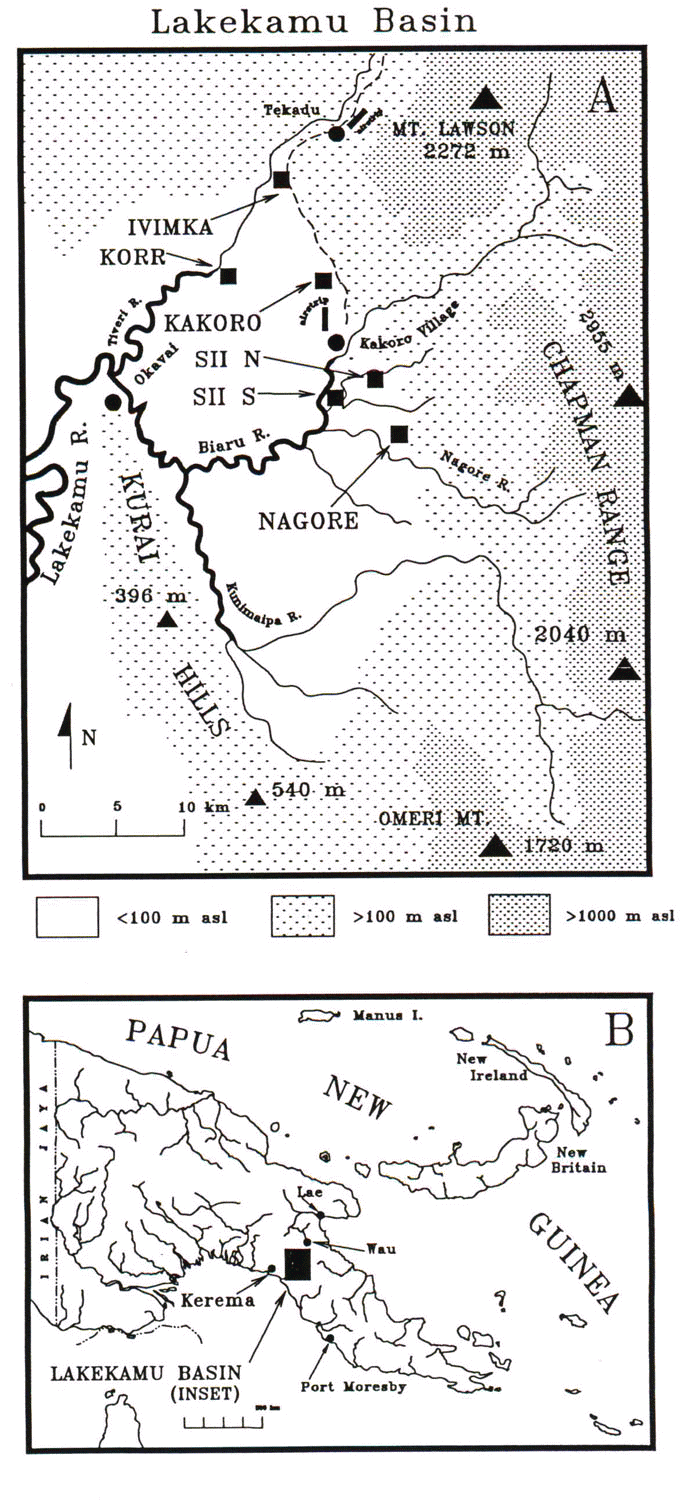
Fig. 2. Species accumulation curve for Basin field surveys from 1979-1998.
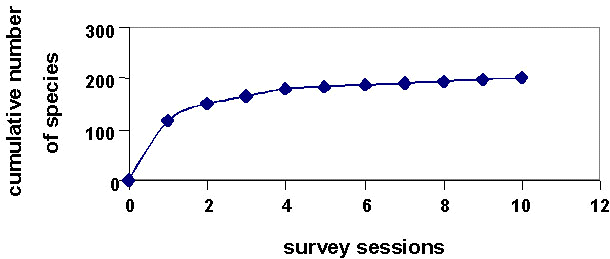
Fig. 3. Survey results vs. length of field season.
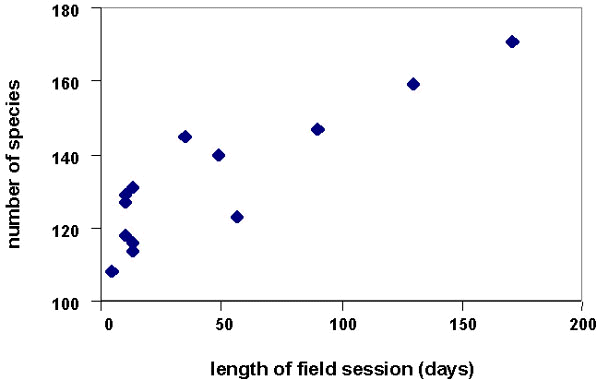
Fig. 4. Species occurrence between field sites.
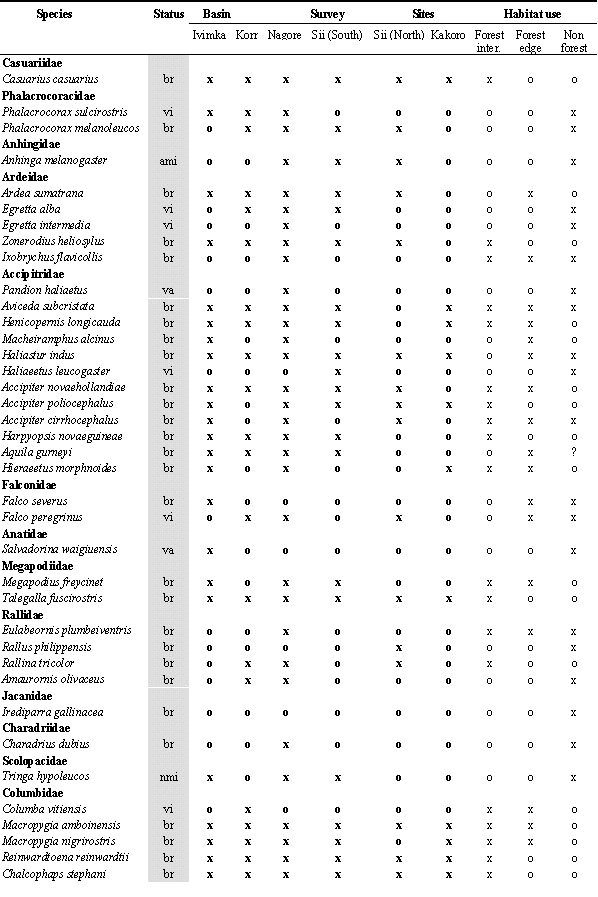
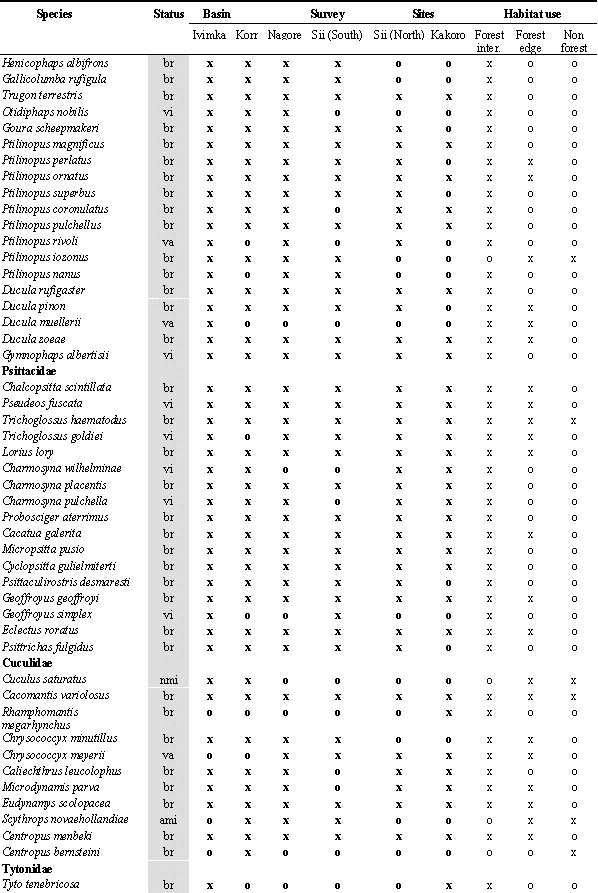
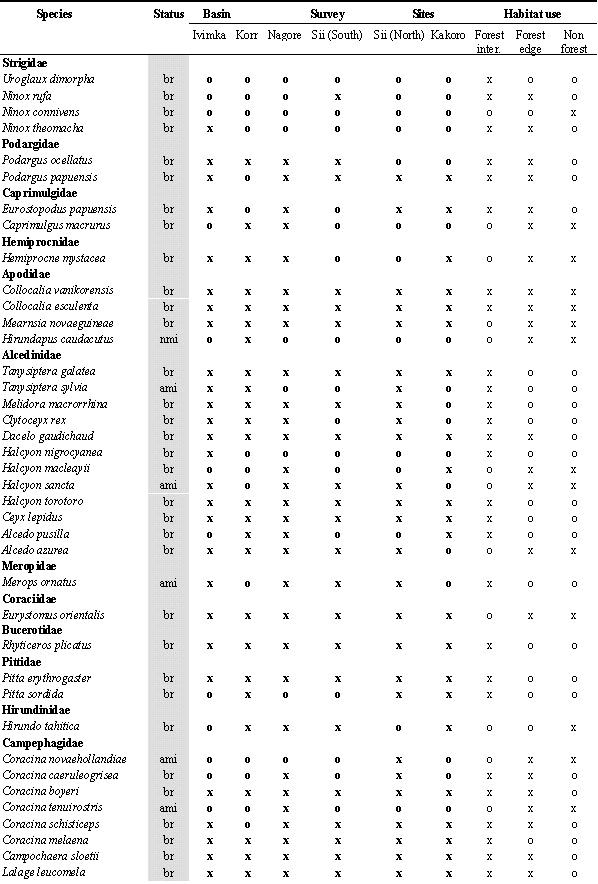
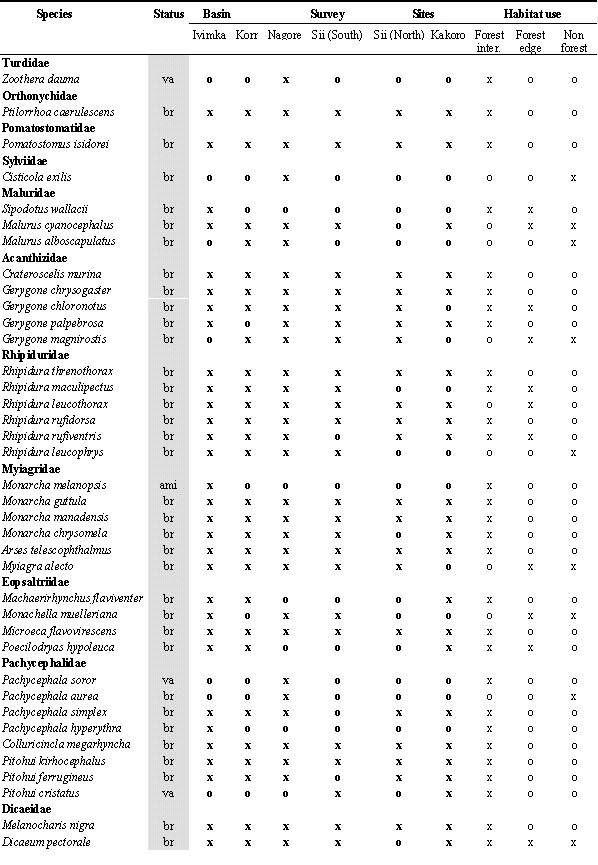
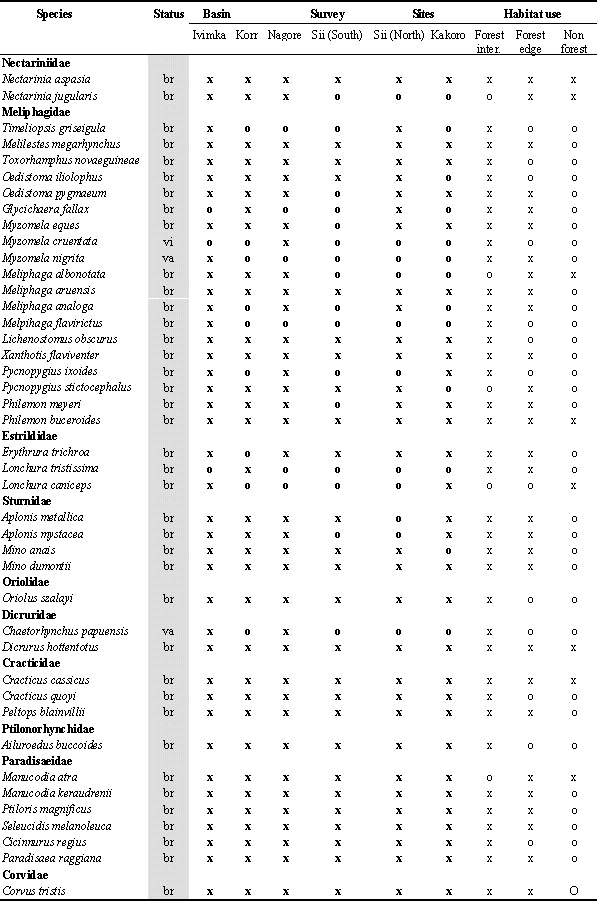
Legend for Abbreviations: br = breeder and resident; vi=local visitor; va=local vagrant; ami=austral migrant; nmi=northern migrant; All lists: o=absent, x=recorded; Number counts: ; for netting = total number of individuals netted in survey (not including retraps) for Ivimka 98 sight surveys = number of days; out of 10 days species was recorded