S43.1: Effects of sampling scale and method on density estimates of tropical birds in Malaysia
Mohammed Zakaria bin Hussin1 & Charles M. Francis2
1Faculty of Forestry, Universiti Pertanian Malaysia, 43400 Serdang, Malaysia, e-mail mzakaria@forr.upm.edu.my; 2Bird Studies Canada, PO Box 160, Port Rowan, Ontario, N0E 1M0, Canada, fax 519 586 3532, e-mail cfrancis@bsc-eoc.org
Zakaria, M. & Francis, C.M. 1999. Effects of sampling scale and method on density estimates of tropical birds in Malaysia. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2554-2568. Johannesburg: BirdLife South Africa.Reliable estimates of population densities of tropical birds are required for many purposes, including understanding the evolution of tropical bird communities, planning reserves, and assessing the effects of land use activities such as selective logging. The few published estimates of population densities of Malaysian birds have been based upon relatively small study plots that do not necessarily provide reliable estimates. We compared these earlier published estimates with density estimates derived from much larger plots using both variable-distance line transect surveys and mark-recapture methods from mist-netting data. For a suite of relatively common babblers at various study areas in Malaysia, we found considerable variation among methods in estimates of density or relative density. Density estimates based upon line transects were higher than those from capture-recapture data, despite expectations that the latter may be biased high. Density estimates based on mist-netting in larger plots tended to be lower than those based on small plots. Variation within plots was of similar magnitude to variation among years, suggesting relative homogeneity of the plots. Measures of the changes in bird communities associated with logging differed between line transect and mist net capture data. Further research, using additional sampling methods, is required to determine which, if any, of these methods produces reliable estimates of density. These studies should be based on large plots (³ 100 ha), both to minimise bias associated with small plots, and to obtain adequate sample sizes for some of the less common species.
INTRODUCTION
Tropical forests support the most diverse communities of birds in the world. To understand the evolution of this tremendous diversity, it is necessary to assess not only the number of species in each area, but also the densities of each and their distribution within each habitat in an area.
Conservation of these communities also requires information on population densities to determine the size of reserves that are required to support viable populations of even the scarcest species (Soulé 1987). Many tropical countries, including Malaysia, were once covered almost entirely by tropical rain forest, but substantial portions of the forest have recently been cleared or selectively logged. Some areas have been set aside as protected areas (parks and wildlife reserves), but there is a need to assess their adequacy. Reserves that are too small are likely to face the selective extinction of many species (Webb et al. 1985, Newmark 1991, Stouffer & Bierregaard 1995, Bierregaard & Stouffer 1997). To minimise or avoid such loss of species when planning reserves and alternative land uses, it is important to know the population densities of different species, as well as factors that influence their extinction probabilities, such as survival and recruitment rates (Lande 1987, Karr 1990).
Reliable methods for estimating densities are also required to assess the impact of habitat degradation, particularly that caused by logging, on bird densities. With relatively limited areas of the original primary forest in most tropical countries designated as reserves or parks, the carrying capacity of selectively logged forest may be critical to the conservation of many species. Research on bird communities in logged forest may be helpful to develop revised logging practices that have a reduced impact on the communities. Most studies to examine the effects of logging on bird communities, especially in Malaysia, have only considered the presence or absence of species in the logged forests, or have at best provided indices of relative abundance (e.g., Johns 1986, 1996; Lambert 1992). If these indices are dependent upon the sampling method, and if the detectability of species varies among sites, the results may not be reliable.
Few estimates of population densities are available for most forest bird species in Malaysia (or elsewhere in south-east Asia). Fogden (1972, 1976) estimated densities of birds in a 20 ha plot in Sarawak based on mist-netting, records of singing birds, and observations of colour-ringed birds. He estimated that 11 species of babblers had territories largely contained within the plot, with between two and 10 pairs per species. An additional three species of babblers used the plot, but their territories were centred outside the plot. Wells (1978) estimated numbers of a selection of understorey bird species based on mist net captures within a 15 ha plot at Pasoh Forest Reserve in peninsular Malaysia. For babblers, he recorded 18 species within the plot, half of which had more than 20 individuals, including one species with 55 individuals. In this case, no attempt was made to differentiate floaters or young birds from resident adults.
From these relatively small plots, it may not be appropriate to extrapolate the results to larger areas for several reasons. First, substantial portions of the territories of many species likely extend beyond the boundaries of the plot, such that numbers cannot reliably be converted to densities by dividing by the area of the plot, especially if no attempt is made to map territories. Second, if there is substantial local heterogeneity in the landscape (beta-diversity), a small plot may not be representative of larger areas within the forest. Third, results from a small plot are likely to be imprecise because fewer birds are included.
The potential for error in density estimates based upon small plots is suggested from some studies of Neotropical bird communities. A study on a 2 ha plot in Panama suggested many species had densities ³ 50 pairs per km2 (Karr 1977). In contrast, an intensive survey on a much larger plot (97 ha) in Peru found that many species were present at densities £ 1 pair per km2, and even the most abundant species had only 30 pairs in the plot (Terborgh et al. 1990). Subsequent work on a larger plot in Panama showed that some of these differences were geographical, but confirmed that densities had a tendency to be overestimated in the small plot (Robinson et al. 1999).
One of the more reliable methods for estimating densities of many tropical forest birds is spot-mapping, which involves delineating territories of singing males, and hence estimating the number of resident family groups (Terborgh et al. 1990). Unfortunately, this method is very labour intensive, and may require several person-months, with field-workers skilled at identifying all species by song, to census an adequate area (e.g., 100 ha) in tropical rainforest. Conversion to numbers of individuals also requires estimates of average group size, as well as numbers of non-territorial individuals (floaters). We have not yet been able to obtain adequate data from any sites in Malaysia using these methods.
Instead, we make use of data from intensive mist-netting on 4 large study plots (110 ha), in three separate areas of Malaysia, supplemented with line transect density estimates in one of these areas, to explore several aspects of bird population densities in Malaysia. For this paper, we concentrate on a suite of understorey forest babblers in Malaysia, which collectively are some of the most frequently caught species in the lowland forests of Malaysia. We compare estimates of density based upon capture-recapture analysis and line transect data with previously published estimates from smaller plots, to evaluate the effects of sampling scale on estimates of population densities. We also compare two methods, mist net samples and line transect samples, to estimate differences in the relative abundance of babblers between primary and secondary forest. We also use mist net capture frequencies to assess the importance of variation within plots relative to variation among years or among study areas.
METHODS
Mist-netting
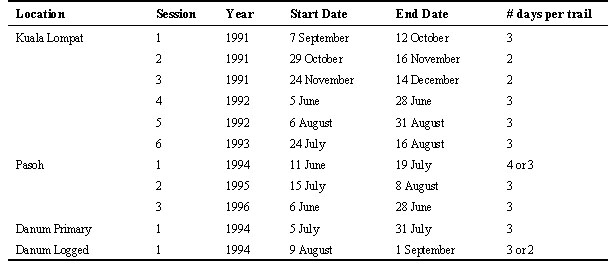
Intensive mist-netting was carried out at four study sites in three different areas in Malaysia. The first site was at Kuala Lompat in the Krau Game Reserve, Pahang, peninsular Malaysia (3° 43'N, 102° 10'E), where a 110 ha grid of trails was established in 1991. The grid consisted of a set of 12 parallel trails running north-south that were 1 km long and 100 m apart. These were joined at the ends and in the middle (i.e. at the 500 m mark) by cross-trails running east-west. The habitat was mature lowland dipterocarp forest, with slightly rolling terrain, and two small streams (one of them seasonal) flowing through the plot.
The second site was at Pasoh Forest Reserve, in the state of Negri Sembilan, peninsular Malaysia (2° 58'N, 102° 17'E). Here, a similar grid of trails was established except that the main set of trails ran east-west, to match better the existing sets of trails in the area, and to minimise disturbance to a botanical plot that overlapped this plot. The 300 x 500 m study plot of Wells (1978) formed the SW corner of this plot. The forest in this area had been more heavily disturbed than the plot at Kuala Lompat, and was of somewhat lower stature. Parts of the plot had been selectively logged in the 1950’s.
The third and fourth sites were around the Danum Valley Conservation Area in the state of Sabah, East Malaysia (4° 57'N, 117° 36'E). The primary forest site was within the conservation area, to the north of the main grid of trails established for general research use in the area. At this site, a similar grid of trails was cut in an east-west direction. The other site was about 5 km east of the primary forest site, in forest that had been selectively logged in 1988, 6 years prior to the survey. Here, a grid was established with the trails running north-south. The terrain in both sites was uneven, with some small streams flowing through, and the Segama River nearby.
During netting sessions at each of the four localities, mist nets, mostly 12 x 2.2 m with 36 mm mesh, were set at approximately 50 m intervals along the whole length of two adjacent 1 km trails and the connecting trails between them. Nets were opened daily, shortly after dawn, checked every one to two hours throughout the day, and closed late afternoon or early evening, before dusk. Nets were usually operated for three days in a row (two days on some sessions) and then moved to the next set of trails on the following day. An entire grid could be covered in six periods of three to four days each (including one day to move nets), although rest periods or other interruptions sometimes extended the sampling period (Table 1). All captured birds were ringed (banded) with uniquely numbered rings, and most individuals also received colour-rings in combinations intended to be unique for each individual of that species at each site. If sufficient trained personnel were available, birds were ringed and released at the nets immediately after capture; otherwise they were brought back to a central point for processing and released there.
At each session, an attempt was made to cover the whole grid with equal effort, but this was not always possible. A major storm in the logged forest grid at Danum, shortly after the trails were cut, caused numerous tree-falls, blocking many of the trails. Because it was not possible to clear most of these prior to netting, and because of additional disturbances due to a herd of elephants passing through, total netting effort was only about half of that in the primary forest grid at Danum. At Pasoh, another storm caused tree falls that blocked about 40% of two of the 12 trails in 1995. One of these, but not the other, was cleared prior to netting in 1996.
Line Transects
Line transect surveys were carried out only at the Danum Valley Forest Reserve (Zakaria 1994). The primary forest surveys were carried out along 4 km of trails, adjacent to or overlapping the plot used for the mist-netting study. These were run in four segments of 1 km each, and were oriented either east-west or north-south. The logged forest surveys were in a different plot from the mist-netting, about 1.5 km north-east of the primary forest plot, in a tract that had been selectively logged with tractors in 1989. Here, the transects were run along four trails each 1 km long, three of them running north-south, and one of them running south-east to north-west. The same trails were surveyed twice per month from November 1990 to February 1992. The first surveys for every line in each month were done in the morning ( starting at 0700) and the second in the late afternoon (starting at 1500). The walking speed was about 500 m per hour. The transects were selected to cover a range of habitats in each site, including edges, small streams, ridges and slopes. All birds heard or seen were recorded, but only birds seen were included in the analysis. For each of these, group size and perpendicular distance to the trail were estimated, the latter in the categories: 0-10 m, 10-20 m, 20-30 m, 30-50 m, > 50 m.
Data Analysis
For analysis, we selected all babblers that regularly use the understorey of the forest, were frequently detected or captured at one or more sites, and were known to occur at all sites (although not necessarily detected on every survey). These species were selected because they include many of the most frequently captured or seen among all species of understorey birds in the forest. As a result, they provide the most reliable data for statistical analysis.
For the mist net data, for species with sufficient captures and recaptures, we used open-population Jolly-Seber capture-recapture models (Pollock et al. 1990) to estimate the size of the population being sampled (N). We included known immature birds as well as adults, for comparison with most of the other survey methods that did not discriminate ages. This was only possible for the two sites (Pasoh and Kuala Lompat) with multiple capture sessions. We then converted these to densities using the simplistic approach of dividing the estimated population size by the area enclosed by the plots (110 ha). We recognise that this is likely to over-estimate densities because some individuals in the sampled population have territories that extend beyond the boundaries, such that the actual effective study area is probably larger than 110 ha. However, the extent of this problem is likely to differ among species, and we do not have any reliable estimates for territory sizes, so we could not make a correction. The implications of this limitation will be considered in the discussion.
Bird densities from line transect data were estimated using program DISTANCE (Buckland et al. 1993). All of the babbler species we considered were regularly seen in groups or in pairs. Species usually in larger groups included Scaly-crowned, Moustached, Sooty-capped, Rufous-crowned, Chestnut-winged, Chestnut-rumped, and Fluffy-backed Tit Babblers (Table 2 for scientific names). Species mostly in pairs included Short-tailed, Horsfield’s, Striped Wren, Black-capped, Ferruginous and Striped Tit Babblers. For birds in groups, the mean distance from the trail to the individual in the middle of the group was estimated, while for pairs, the mean distance to the first individual seen was recorded. Density estimates were then calculated for groups, and multiplied by mean group size to obtain the density of individuals.
For comparison, we also extracted density estimates for the same suite of species from Fogden (1976) for Semengo Forest Reserve, near Kuching in Sarawak, and from Wells (1978) for a 15 ha subset of the current mist-netting plot at Pasoh. Fogden (1976) presented his estimates as the number of resident adults of each species present in a 20 ha plot at Semengo in December 1975, based on mist-net captures and observations of colour-ringed individuals in the plot during intensive study throughout that month. He included only pairs for which he judged at least 70% of their territory was within the plot. We converted these estimates to numbers of individuals per 100 ha by multiplying them by 5. Wells (1978) estimated the number of birds in a 15 ha plot based on the number of different individuals captured between February and September 1973, assuming that most birds were resident, in family groups in the site, during this period (which is the main breeding season). As with our own mist-net data, we converted these to densities using the simplistic assumption that the area they were using was only the 15 ha plot. On this basis, we converted these to numbers per 100 ha by multiplying by 6.7.
To compare sampling methods for estimating changes in bird communities in response to logging, we used line transect and mist net data from around Danum Valley in Sabah. The line transect data provided estimates of actual densities, but for the mist-netting data we were only able to obtain indices of relative abundance of species because we only had a single netting session in each 110 ha plot at Danum. Furthermore, even to obtain indices, we had to double the counts in the logged forest plot to take into account the reduced netting effort (50%) in that plot. Thus, we had to base this comparison on changes in relative abundance.
Finally, we tested whether small scale geographic variation or heterogeneity (i.e. within a 110 ha plot) could have a substantial effect on density estimates. To do this, we tallied separately captures within each half of a 110 ha plot at the primary forest sites at Kuala Lompat, Pasoh, and Danum (the logged forest site at Danum was excluded because of uneven coverage). The grids were divided in the middle of each trail (birds on the midline were excluded) to avoid temporal variation associated with covering different trails on different dates. Each bird was considered only on its first capture during a session in a particular half of the netting grid. For these comparisons, we only considered the first survey each calendar year at Kuala Lompat. Again, we based this comparison on relative abundance, because a single capture session does not provide adequate data for estimating densities. We measured similarities in relative abundance between the two halves of each netting grid, and compared them with similarities across years within each half grid, as well as similarities among study areas. We also calculated similarities among years and among study areas using data from the whole of each grid.
RESULTS AND DISCUSSION
Population Densities
We compared estimates of the total numbers of individuals per 100 ha derived from our capture-recapture analyses and line transect surveys with published estimates of densities for these same species from Fogden (1976) and Wells (1978) (Table 2).
Only the estimates from Pasoh refer to the same location for two different studies (both plots overlapped), although they were 20 years apart. In this case, with the exception of Sooty-capped and Horsfield’s Babblers, the capture-recapture estimates from the large plot (our study) indicate much lower densities than those from the small plot (Wells 1978). This discrepancy could be due partly to bias associated with the simplistic assumption that the area enclosed by the netting areas represented the size of the study area. As noted earlier, and recognised by Wells (1978), many birds have territories that extend beyond the boundaries of the netting area. The bias caused by this problem is dependent upon the size of the study area. For example, if we assume an effective catching area of an additional 100 m around the outside of the plot, the 300 x 500 m plot of Wells (1978) becomes approximately 500 x 700 m or 35 ha rather than 15 ha—a 2.3-fold increase. The 110 ha plot becomes 156 ha which is only a 1.4-fold increase. Thus, unless reliable estimates of the extent of the additional territory used by the birds can be obtained (see below), extrapolations from population estimates for finite plots within more extensive contiguous habitat will over-estimate densities much more strongly for small than for large plots. Furthermore, even if estimates of this extra territory are available, uncertainty in those estimates will lead to much greater error for small plots than large plots.
Methods for estimating the extent of the bias associated with unclosed boundaries, and hence correcting for it, have been developed for closed-population capture-recapture models by using nested plots of different sizes (Otis et al. 1978), but we do not have adequate data to use those methods. Estimating the extra area based on behaviour is possible, but complex, because some birds caught in the grid are likely to range much farther than 100 m from the net lines, while other birds may have home ranges close to, but not overlapping, the study area. The correction will necessarily vary among species.
Open population capture-recapture models have limitations for estimating bird populations based on mist-netting data, because their assumptions are not perfectly met. Some of the most critical assumptions are that all individuals have equal capture probabilities and that all emigration from the area is permanent. If marked individuals have lower capture probabilities than unmarked individuals (as could result from net shyness), densities are likely to be overestimated. Nevertheless, these should still provide an improvement over simple capture totals. For most species, the expected bias in capture-recapture estimates (upwards) was in the opposite direction to the observed differences from Wells (1978) study. Simple capture totals will under-estimate relative abundance of species with lower capture probabilities (e.g. because they often forage higher in the canopy). Also, in Wells (1978) study, the totals included all birds captured over an 8-month period. Even if there were little turn-over in the breeding adults during this period, turn-over in transients or young birds could mean that only a portion of the birds recorded were actually present at any given time in the plot.
The Sooty-capped Babbler provides an exception to the pattern of lower density estimates from the larger plot. This species was rarely captured by Wells (1978). We could not obtain a reliable density estimate for this species from capture-recapture data because, although many birds were present (about 30 individuals were captured per year), very few were recaptured (of the 30 individuals captured the first year, 3 were captured in the second year, and 3 different individuals in the third year). This potentially suggests an extremely large population (>500 birds), but it could also be due to considerable heterogeneity in capture probabilities (e.g. marked individuals may avoid subsequent capture), or movement of individuals in and out of the study area. Both of these factors could lead to over-estimate of populations. In any case, even based on numbers captured, this species was clearly more abundant than reported by Wells (1978). This species tends to forage in the canopy or in forest gaps caused by treefalls or other disturbances. Changes in the structure of the forest over the 20 years between the studies may have led to increased abundance of this species, or an increase in its appearance in the understorey. There may also be differences between the 15 ha plot and the remainder of the 110 ha plot, although this species was regularly observed in the original plot during the recent studies (C. M. Francis, unpublished data).
Interpreting differences among estimates from the other studies in Table 2 is complicated by the fact that geographic variation is confounded with differences in estimation techniques. However, we can obtain a crude estimate of geographic variation in average abundance of these species by comparing the numbers captured with similar netting effort (3-days per net line) during netting sessions at each of the 3 primary forest study sites during June/July, the time of year when capture rates appeared to be highest. The total for all of the study species of babblers combined was 336 at Danum, 357 at Kuala Lompat, and 292-294 at Pasoh. Especially considering that the Pasoh estimates were based on 5-10% less effort (due to blocked trails), these results suggest broadly similar average densities of these 14 species of babblers at the three sites.
Density estimates based upon capture-recapture (Table 2) were similar in magnitude to those obtained by the "census" of individuals in a 20 ha plot at Semengo (Fogden 1976). This was despite the fact that the capture-recapture estimates are likely to over-estimate densities of many species (due to uncertainty in the sample area, as well as bias associated with net shyness). Also, the netting totals include floaters and immature birds (though the latter were rarely >20% of the catch), while Fogden (1976) considered only breeding pairs.
In contrast, density estimates from the line transect surveys were generally much higher than from either of these methods. Without knowledge of the actual population densities at each site, it is not possible to tell which method produces the least biased estimates. Nevertheless, if we are correct in suspecting that capture-recapture models are over-estimating densities, then the transect surveys must be even more strongly biased. One possible source of bias could be due to movement of bird flocks relative to the observer, but this needs to be explored more thoroughly with additional field work.
Effects of logging
At Danum, only the line transect surveys provide estimates of actual densities of various babbler species in pristine and in logged forest. However, the mist net data from each plot can provide indices of relative abundance at each site, after correcting for effort (Table 3). Thus we based our comparisons on the relative abundance of species from each method. We only analysed 12 of the 14 babbler species, because density estimates were not calculated from transect data for two species.
Based on the line transect data, all species except the Fluffy-backed Tit-Babbler were substantially more abundant in primary than logged forest. The mist-net results agreed in showing that Fluffy-backed Tit-Babbler was more abundant in logged forest, but they also indicated similar or higher densities in logged forest for at least 3 other species. To determine whether both methods were consistent in estimating the relative abundance of each species in the two habitats, we calculated Pearson correlation coefficients between density indices for each of the 12 species from both methods as measures of similarity between samples. The correlation between estimates of relative abundance in primary forest from each method (r = 0.60, P = 0.04) was higher than the correlations across habitats (transect in primary versus captures in logged, r = 0.14, n.s.; captures in primary versus transect in logged, r = 0.45, n.s.), but the lowest correlation was between methods in logged forest (r = -0.07, n.s.). The highest correlations were between habitats using the same methods (transects: r = 0.93, P < 0.0001; captures: r = 0.56, P = 0.06). This implies that differences in detectability among species with each method were greater than differences in density between habitats. Different conclusions would be reached about which species were most strongly affected by logging depending upon which sampling method was used.
Effects of Time and Spatial Heterogeneity
One of the potential disadvantages of a small plot is that, if there is extensive heterogeneity within a habitat, species present in an overall study area may be missed in that particular plot. For example, Beehler (1999) found considerable patchiness in the distribution of several common bird species in lowland forest in New Guinea even over relatively short distances.
To test for small scale patchiness, we compared the two halves of the study plots in primary forest at Danum, Kuala Lompat, and Pasoh. At all 3 sites, both halves were consistent in recording the presence or absence of each babbler species (Table 4), although some of the less common species at Kuala Lompat and Pasoh were missed in one or the other half in some years.
We used simple Pearson correlation coefficients as an index of similarity between the two halves of the study areas at each site. Within years (averaging across the three years available for Kuala Lompat and Pasoh), similarities between halves ranged from moderate at Pasoh to very high at Danum (Table 5). For those sites with multiple years of data, these were similar to, or slightly higher than similarities among years within halves (Table 5). This suggests either that small scale geographic variation is similar in magnitude to annual variation, or that most of the variation was due to sampling variation (not all birds are caught in each session). In contrast, there were few similarities in the relative abundance of species among sites.
We also calculated indices of similarity among years and among plots for annual species totals based on the whole of each plot, including the secondary forest plot at Danum (Table 6). In this case, the average correlations between years were somewhat higher than those based upon the half-plots. This is to be expected if much of the variation was due to sampling error because the larger plots, with more individuals, would have less sampling error. There was again very little similarity among study areas, except in the comparison of the primary forest and logged forest sites at Danum, which were nearly as similar to each other as the two halves of the study plots at Pasoh or Kuala Lompat (Table 5).
CONCLUSIONS
Our conclusions from these analyses are necessarily less rigorous than they might have been had the data been collected specifically to evaluate methods for estimating density. Nevertheless, in view of the high cost and effort required to gather data of this nature, we felt it appropriate to make use, as much as possible, of existing data sets. As a minimum, these analyses can help to guide future research.
Even allowing for annual and geographic variation, we found considerable variation among methods in the density estimates, with some methods suggesting much higher densities than others. Furthermore, in the studies at Danum, different conclusions were reached about which species were most strongly affected by logging, depending upon which methods were used.
Although two of these methods (line transect sampling and capture-recapture analysis) are based upon sound statistical theory, neither will produce reliable estimates if its assumptions are violated (Pollock et al. 1990, Buckland et al. 1993). Lower capture probabilities of already marked birds (if they learn to avoid nets among sessions) as well as under-estimates of the area being sampled will both lead to over-estimates of density from capture-recapture data. The fact that density estimates from line transects were even higher, suggests they may also be over-estimated, perhaps because of difficulties in judging distances, or violation of the assumption that animals are moving only slowly relative to movement of the observer (Buckland et al. 1993).
Further research using additional sampling methods is required to determine which, if any, of these methods produces reliable estimates of density. Spot-mapping of singing males, preferably combined with following and observing colour-marked individuals on a large plot (110 ha) might produce the most reliable estimates of densities of territorial birds (Robinson et al. 1999), although this does not take into account non-resident individuals or floaters. This is similar to the approach used by Fogden (1976), but he used a relatively small plot, which may have increased error in his estimates. Unfortunately, gathering such data from large plots can require several months with highly skilled labour (Terborgh et al. 1990). Although we attempted to gather such data at one site in the current study, we did not have sufficient time or skilled personnel to complete the surveys. We are currently exploring the possibility of using new technology (directional arrays of microphones) to map the locations of singing birds, although it is not yet clear how well that will work in the dense vegetation of tropical forests.
With respect to effects of plot size, we did not find consistent results in terms of bias in density estimates. For methods based on mist-net captures a larger plot was clearly preferable, but estimates based on spot-mapping in a 20 ha plot (Fogden 1976) that attempted to take into account the fact that some territories extended outside the plot, did not appear to be strongly biased. Furthermore, we found little evidence for small scale variation in densities within our study plots. However, this last result must be treated somewhat cautiously, because it was based only on the more abundant babblers in the forest. These babblers may be more habitat generalists than other bird species. Further studies, providing density estimates for all of the species in the bird community, are required to measure geographic variation more effectively. Nevertheless, even if there is little heterogeneity within plots, density estimates should be based on as large a plot as practical (³ 100 ha), both to minimise bias due to edge effects, and to obtain adequate sample sizes, especially for less common species.
From the perspective of conservation, the critical species will be the rarer species, for which the largest areas of habitat will be required to ensure their protection (Thiollay 1989). Unfortunately, for the rarest species, even with a large plot, we doubt that it will be possible to obtain sufficient data from any of these methods to produce statistically reliable estimates of density. One possible alternative is an indirect approach, first estimating density of common species, and then extrapolating these to rarer species based on estimates (e.g. through mist net captures or observational surveys) of the relative abundance of the rarer species compared to the more common species.
To evaluate the effects of habitat disturbance or the quality of individual patches of forest, it is important to assess not only population density, but also population viability. Unless the forest patch is completely isolated from other patches, such that no immigration or emigration can occur, a population could be present in an area, but not successfully reproducing (a population "sink"). It could be sustaining itself only through recruitment from other areas ("sources"). This was found to be the case for many forest birds in isolated forest fragments in certain parts of North America (Robinson et al. 1995). If similar dynamics occur in tropical communities, populations in some areas could disappear if those areas become isolated from their sources. Evaluation of source/sink dynamics requires methods for assessing demographic parameters (survival and recruitment rates) in addition to those for assessing density.
Although obtaining adequate information on densities and population dynamics of tropical forest birds is difficult and time consuming, such information is essential to assess properly the status of bird populations in tropical countries such as Malaysia. Only with this information will it be possible to determine whether the bird communities are adequately protected for future generations, and to take appropriate actions if they are not.
ACKNOWLEDGMENTS
We thank the Economic Planning Unit of the Malaysian Prime Minister’s Department, the Sabah Foundation, the Sabah Wildlife Department, the Forest Research Institute of Malaysia (FRIM) and the National Parks and Wildlife Department (PERHILITAN) for permission to carry out research on birds in Malaysia. The Danum Valley Rainforest Research and Training Programme, the Royal Society, the Wildlife Conservation Society, Bird Studies Canada, Universiti Malaya, Universiti Pertanian Malaysia, and Universiti Kebangsaan Malaysia all provided financial or logistic support. We thank Prof. Nordin Haji Hassan, Prof. Yong Hoi Sen, and Dr. David R. Wells for advice, ideas, and support.
REFERENCES
Beehler, B.M. 1999. Patchiness of bird distributions in a lowland forest in New Guinea. In: Adams, N.J. & Slotow, R.H. (eds) Proc.22 Int. Ornithol. Congr., Durban: this volume.
Bierregaard, Jr., R.O. & Stouffer, P. 1997. Birds in forest fragments. In: Laurence, W.F. & Bierregaard, R.O. (eds) Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities. Chicago, Illinois; University of Chicago Press: 138-155.
Buckland, S.T., Anderson, D.R., Burnham, K.P. & Laake, J.L. 1993. Distance Sampling: estimating abundance of biological populations. Chapman & Hall, London.
Fogden, M.P.L. 1972. The seasonality and population dynamics of equatorial forest birds in Sarawak. Ibis 114: 307-343.
Fogden, M.P.L. 1976. A census of a bird community in tropical rain forest in Sarawak. Sarawak Museum Journal 24: 251-267.
Johns, A.D. 1986. Effects of selective logging on the ecological organisation of a peninsular Malaysian rain forest avifauna. Forktail 1:65-79.
Johns, A.G. 1996. Bird population persistence in Sabahan logging concessions. Biological Conservation 75:3-10.
Karr, J.R. 1977. Ecological correlates of rarity in a tropical forest bird community. Auk 94:240-247.
Karr, J.R. 1990. Avian survival rates and the extinction process on Barro Colorado Island, Panama. Conservation Biology 4:391-397.
Lambert, F.R. 1992. The consequences of selective logging for Bornean lowland forest birds. Philosophical Transactions of the Royal Society, London B 335: 443-457.
Lande, R. 1987. Extinction thresholds in demographic models of territorial populations. American Naturalist 130: 624-635.
Newmark, W.D. 1991. Tropical forest fragmentation and the local extinction of understory birds in the eastern Usambara mountains, Tanzania. Conservation Biology 5: 67-77.
Otis, D.L., Burnham, K.P., White, G.C. & Anderson, D.R. 1978. Statistical inference from capture data on closed animal populations. Wildlife Monographs 62: 1-135.
Pollock, K.H., Nichols, J.D., Brownie, C. & Hines, J.E. 1990. Statistical inference for capture-recapture experiments. Wildlife Monographs 107: 1-97.
Robinson, S.K., Thompson II, F.R., Donovan, T.M., Whitehead, D.R. & Faaborg, J. 1995. Regional forest fragmentation and the nesting success of migratory birds. Science 267: 1987-1989.
Robinson, W.D., Brawn, J.D., Terborgh, J. & Robinson, S.K. 1999. Structure of a lowland Panama bird community: effects of spatial scale and biogeographic history. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2591-2606. Johannesburg: BirdLife South Africa.
Soulé, M.E. 1987. (ed.) Viable populations for conservation. Cambridge Univ. Press.
Stouffer, P.C. & Bierregaard, Jr., R.O. 1995. Use of Amazonian forest fragments by understory insectivorous birds. Ecology 76:2429-2443.
Terborgh, J., Robinson, S.K., Parker III, T.A., Munn, C.A. & Pierpont, N. 1990. Structure and organization of an Amazonian forest bird community. Ecological Monographs 60: 213-238.
Thiollay, J.-M. 1989. Area requirements for the conservation of rain forest raptors and game birds in French Guiana. Conservation Biology 3:128-137.
Webb, L.J., Hopkins, M.S., Young, P.A.R., Kikkawa, J. & Lovejoy, T.E. 1985. Conservation of tropical rain forest isolates. In J. Davidson, J., Tho Yow Pong, & Bijleveld M. (eds) The Future of Tropical Rain Forests in South East Asia. International Union for Conservation of Nature and Natural Resources, Switzerland: 55-68.
Wells, D.R. 1978. Numbers and biomass of insectivorous birds in the understorey of rain forest at Pasoh Forest. Malay. Nat. J. 30: 353-362.
Zakaria, M. 1994. Ecological effects of selective logging in a lowland dipterocarp forest on avifauna, with special reference to frugivorous birds. PhD. thesis, Universiti Kebangsaan Malaysia.
Table 1. Dates of netting sessions at each of the four study sites.
Table 2. Estimates of densities (individuals per 100 ha) of selected babblers from various methods in 5 different areas of lowland rainforest in Malaysia.
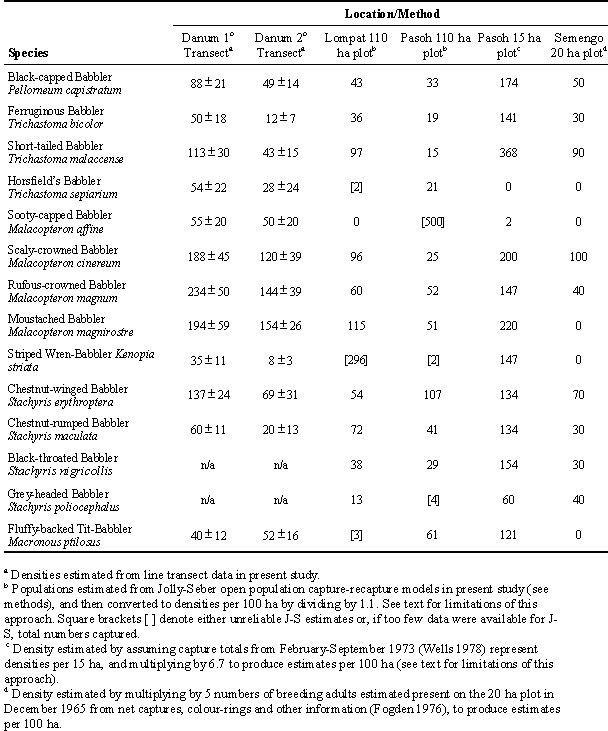
Table 3. Comparison of indices of relative abundance of 12 species of babblers in primary forest and logged forest at Danum Valley Forest Reserve, Sabah, as estimated by line transect data, or mist-net capture data. Net captures in the secondary forest were doubled to correct for about 50% less netting effort at that site.
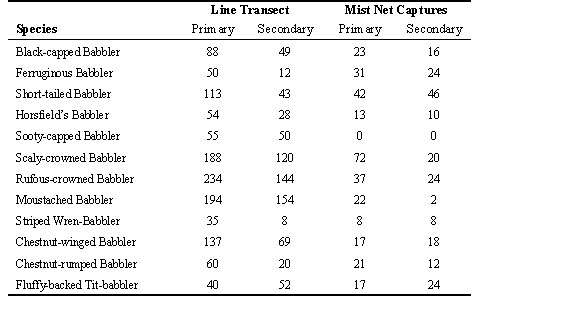
Table 4. Number of individuals of each species of babbler captured on each half of the study grids during the first netting session in each year at Danum Valley (primary forest only), Kuala Lompat and Pasoh.
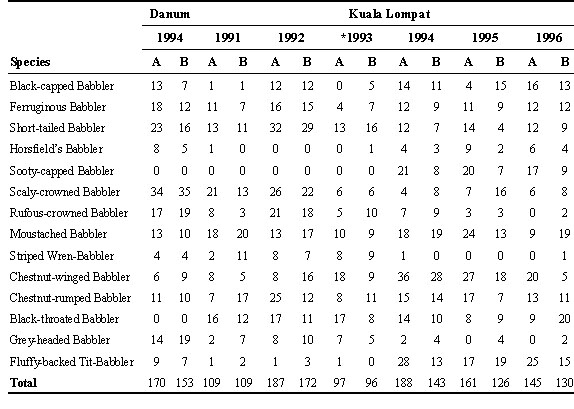
Table 5. Indices of similarity between the two halves of each study grid, between pairs of years, and between sites. All comparisons are based on capture totals in each half of a study area in any given session (only the first session in a year is included). Indices represent correlations between capture totals in mist nets of each of the 14 babbler species. If multiple comparisons were possible, the average of all such correlations is presented (e.g., the comparisons between halves within a year for Kuala Lompat or Pasoh represent the means of 3 years each). Significant (P < 0.05) correlations are shown in bold.

Table 6. Indices of similarity between years and between sites based on complete study grids at each site. Indices represent correlations between mist net capture totals for each of 14 babbler species. If multiple comparisons were possible, the average of all correlations is presented. Significant correlations (P < 0.05) are shown in bold.
