S41.2: Dispersal distance and its demographic consequences in the Florida Scrub-jay
John W. Fitzpatrick1, Glen E. Woolfenden2 & Reed Bowman3
1 Cornell Laboratory of Ornithology, 159 Sapsucker Woods Rd., Ithaca, NY 14850, USA, e-mail
jwf7@cornell.edu; 2,3Archbold Biological Station, PO Box 2057, Lake Placid, FL 33862 USA, e-mail gwoolfenden@archbold-station.org Fitzpatrick, J.W., Woolfenden, G.E. & Bowman, R. 1999. Dispersal distance and its demographic consequences in the Florida Scrub-jay. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2465-2479. Johannesburg: BirdLife South Africa.Demographic monitoring of Florida Scrub-Jays, both in continuous oak scrub habitat and in isolated scrub patches amidst prairie and pine woods, revealed delayed dispersal and extreme philopatry. Absolute dispersal distances were longer in patchy habitat than in continuous, but median number of jay territories traversed was similar. In both areas a few jays, virtually all females, dispersed long distances (maxima: 35 km within continuous habitat, 18 km within patchy habitat). Long-distance dispersal was mostly by young jays that bred at age 2, while jays delaying dispersal beyond age 2 tended to disperse short distances. Annual reproductive success did not differ between short-distance and long-distance dispersers, but survivorship as a breeder and, consequently, lifetime reproductive success was significantly higher among jays dispersing short distances. Surprisingly, males who became breeders through territorial 'budding' showed exceptionally low survivorship during the first two years as breeders. Budding appears to be initially costly, but those individuals that survive the first two years then show very high survival, and their lifetime reproduction is nearly as high as that of other short-distance dispersers. Short-distance dispersal appears to be selectively favoured in the Florida Scrub-Jay, which has led to a dispersal strategy based on monitoring the neighbourhood from the relative safety of the natal territory. The home-based dispersal strategy is pursued even in fragmented landscapes, where fewer territories can be monitored without leaving home. This fact, combined with the high cost of dispersing across unsuitable habitat, has important implications for long-term management of this endangered and rapidly declining species.
INTRODUCTION
Dispersal characteristics vary widely among organisms, and these differences provide grist for numerous questions in comparative life history theory (e.g. Greenwood 1980, Johnson and Gaines 1990). Of special interest in the study of avian social behaviour are the tendencies for some species to delay natal dispersal and to disperse over extremely short distances, while others - even closely related forms - disperse early and move much greater distances. As pointed out by many authors (e.g. Stacey and Ligon 1987, 1991; Koenig et al. 1992), co-operative breeding often is associated with extreme natal philopatry. This association suggests that progress in understanding the evolution of co-operative breeding depends in part on understanding the factors leading to reduced dispersal. Here we examine attributes of dispersal behaviour in a well-studied co-operative breeder that is among the most sedentary known. We describe aspects of dispersal tactics in both continuous and fragmented habitat, and we provide evidence for differences in lifetime fitness between individuals following short-distance versus long-distance dispersals.
The Florida Scrub-Jay Aphelocoma coerulescens is a non-migratory species restricted to relict patches of fire-maintained, stunted oak scrub in the peninsula of Florida (Woolfenden and Fitzpatrick 1984, 1996). Family groups containing a single breeding pair defend large territories (5-15 ha) year round. Virtually all fledged offspring remain on and help defend their natal territory for at least one year. During this period of residency, most of these helpers also deliver food to any nestlings and fledglings produced by the breeding pair. The jays forage on arthropods and small vertebrates during all months and on acorns harvested and cached by the thousands during late summer and fall. Natal dispersal (movement from birthplace to breeding site) typically commences during the second year of life, and often involves dispersal forays among neighbouring territories. Males become breeders primarily by replacing lost breeders in nearby territories or by budding a portion of their natal territory (Woolfenden and Fitzpatrick 1978). Rarely, males may inherit the natal territory outright, and even more rarely they create a new, de novo territory away from their natal one. Females either replace lost breeders or pair with a male that is budding, inheriting, or establishing a new territory de novo. Inheriting and budding by females is extremely rare. Regardless of mode, exclusive ownership of space is a requisite for successful pairing and for breeding. Breeding space competition is extremely intense most years (Fitzpatrick and Woolfenden 1986).
In this paper we address several questions about the nature and aftermath of natal dispersal in the Florida Scrub-Jay. (1) How do dispersal distances of jay populations living in a highly fragmented landscape compare with those in continuous habitat? (2) Given that most individuals engage only in short-distance dispersal, what features, if any, characterise those that disperse the farthest? Are these individuals that tried and failed at shorter-distance movements, or are they young birds engaging in an explicit strategy of dispersal as opposed to remaining home, as in Red-cockaded Woodpeckers Picoides borealis (Walters 1990, Walters et al. 1992a). (3) Do reproductive success and annual survival of breeders vary with their natal dispersal distance, and, if so, does this variation help explain why long-distance dispersal is rare in this species?
METHODS
Florida Scrub-Jays are unusually easy to observe and study because they live in a low-growing, open, shrub habitat, and because they can become remarkably tame to humans with only a little effort at offering them food. From 1969 through the present we have studied intensively a population of such tamed jays at Archbold Biological Station in south-central peninsular Florida (Highlands County). Our main study tract ('Demography Tract') slowly expanded during the study period from about 150 ha in the 1970s, to about 450 ha between 1988 and the present. The number of jay families encompassed in this focal area grew from about 20 in 1970 to about 60 by 1988, and has remained approximately stable since that year. Throughout this study tract we have colour-ringed all resident scrub-jays, both natal and immigrant. We monitored all nesting activity each spring, and we conducted complete censuses each month of all family groups and individuals in the tract.
Crucial to the present paper are several facts derived from the 317 consecutive monthly censuses we have conducted since April, 1971 (see also Woolfenden and Fitzpatrick 1984). Florida Scrub-Jays live in closely knit and easily censused family groups, hence, we rarely fail to locate a jay that is actually present in the tract. Absence of a jay from a monthly census represents either a death or a dispersal (temporary or permanent) away from our study tract. At Archbold, jays essentially never move from their territory once they become breeders. Therefore, absence of a breeder represents a death. The same is true, with rare exceptions (see Results), for jays in their first year of life. However, for non-breeding jays older than one year we are unable to distinguish a death from a dispersal event, in the absence of corroborating evidence of death or later sightings away from the study tract. Fortunately, we do have considerable opportunity to locate such dispersers.
Over 30 years we have accumulated extensive information about the frequency and distribution of successful dispersers that moved outside our Demography Tract. Every few years between 1975 and 1991 we conducted extensive searches for marked dispersers by censusing otherwise unstudied jay families throughout Highlands County, up to 35 km from our study tract. Such efforts were especially thorough, involving several hundred jay families, in 1979, 1985, and 1991. In 1992-93 we co-ordinated a statewide survey of Florida Scrub-Jays to produce a detailed distribution map for the species as a whole (Stith et al. 1996). Approximately 3,200 of the 4,000 remaining family groups were directly observed by biologists during that 18-month period (Fitzpatrick et al. 1993). In 1997, as part of RB’s study of suburban jay demography, about 300 jay families throughout Highlands County were censused in order to colour-ring the juveniles.
Supplementing our explicit efforts to locate dispersers are additional, intensive studies of colour-ringed jays elsewhere in Highlands County. These began in 1986 on a comparably sized study tract immediately adjacent to our own (e.g. Mumme 1992), and in 1991 by RB on a suburban study tract about 10 km from our own. Those studies incorporated regular searches for marked jays in areas adjacent to all three sites, and provided both opportunity to locate dispersers from the Demography Tract and sources of a few colour-ringed immigrants into the tract.
All the Highlands County study tracts are located near the southern end of the 'Lake Wales Ridge,' a sandy, north-south oriented, ancient shoreline deposit originally supporting Florida’s most extensive and diverse oak scrub communities (e.g. Christman and Judd 1990). About 50 km northeast of Archbold Biological Station lies a separate, smaller scrub ridge at the Avon Park Air Force Range (APAFR), located in both Highlands and Polk counties. Here a population of scrub-jays has been marked and studied by RB since 1992. A major difference between the study tracts on these two ridges is the configuration of habitat acceptable to Florida Scrub-Jays: at Archbold, on the Lake Wales Ridge, the scrub is expansive and mostly continuous; at APAFR the scrub mostly consists of small patches (2 - 300 ha) interspersed with natural habitats (mainly pine forest or prairie) unacceptable to the jays. Habitat patches are separated from one another by distances of 50 m to 10 km, with most gaps falling between 0.1 and 1.0 km. Suitable habitat constitutes only about 10% of the total habitat in the study area. This difference in habitat configuration between the two study areas has allowed us to study the effect of habitat fragmentation on Florida Scrub-Jay dispersal distances.
Except for comparisons of dispersal distance between fragmented and unfragmented habitat, data we present in this paper are from the long-term study on the Demography Tract at Archbold Biological Station. Natal dispersal distances are summarised through the 1996 breeding season. Only jays that became paired on a territory and tended at least one egg were considered to have become breeders, and therefore to have completed a dispersal event. Dispersal distances were counted (for territories traversed) and measured (linear distance) by hand on territory maps generated annually for both study areas. Individuals that inherited or budded their territories were considered to have dispersed 0 and 0.5 territories, respectively. Only 1 female inherited her natal territory, and 2 females budded. These three females were lumped with those dispersing 1 territory in all analyses of reproduction and survival with dispersal distance. Movement by an established breeder to a new territory ('breeding dispersal') is rare at Archbold, and is ignored in this paper. We limited our analyses of annual reproduction and survival to those jays for which we had complete post-dispersal records; this excluded a few jays that dispersed to the periphery of our expanding study tract, and for which we lacked certain relevant data. To analyse lifetime reproductive success with dispersal distance, we considered only jays that became breeders in our study tract between 1971 and 1991, and for which we have complete lifetime breeding records (N=140 females, 127 males). Of these cohorts, only 4 jays remained alive at this writing (June 1998; 1 immigrant male, 1 male that budded, plus 1 male and 1 female that dispersed 1 territory away). We treated these 4 jays as if their lives were completed; any additional lifetime output by them will not measurably alter our results.
RESULTS
Dispersal Curves in Continuous and Fragmented Habitat
The frequency distribution of Florida Scrub-Jay natal dispersals is strongly leptokurtic, and the tail of the curve is significantly biased toward females (Fig. 1). Modal dispersal distance for both sexes is only 1 territory, and fully 25% of males inherit at least a portion of their natal territory. Less than half the females, and less than 25% of the males, disperse more than 2 territories from their birthplace. Only among females did a significant number of individuals disperse 10 or more home-range distances from the natal territory (i.e. 'long-distance dispersal' as defined operationally by Shields 1982).
Measured in actual distance, jays living in naturally fragmented habitat dispersed significantly farther than those in continuous habitat (Fig. 2a). In continuous habitat at Archbold, mean dispersal distance was 1593 + 2517 m, and the median was only 680 m. In fragmented habitat at APAFR, the mean was 2956 + 4032 m, and the median was 900 m. This difference disappeared, however, when we measured dispersal distance in terms of home-range diameters, or territory widths, rather than as a metric distance (Fig. 2b). The strikingly similar distributions of dispersal distances when measured in territories traversed show that even in a fragmented landscape, Florida Scrub-Jays exhibit pronounced natal philopatry. Most jays recruit as breeders within the immediate neighbourhood of their birth.
Dispersal Distance and Age at Departure
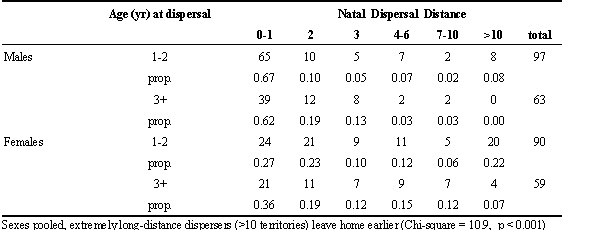
Florida Scrub-Jays become breeders at different ages (Woolfenden and Fitzpatrick 1996). A few jays first breed at age 1, a majority at age 2, and the remainder at ages 3-5. We compared the dispersal distances of jays that first bred by age 2 to those that had delayed breeding at least until age 3 (Table 1). These dispersal distances were similar, with one notable exception. In both males and females, jays that dispersed the longest distances (10 or more territories from home) were significantly biased toward the early dispersers (Chi Square = 10.9, p<.001). A few of these long-distance dispersers even departed their natal territory well before age 1. It is clear, therefore, that some jays chose to depart from their home neighbourhood well before exhausting more local dispersal opportunities. In an attempt to explain these early departures from home, we compared features of family size, family composition, family stability, and neighbourhood density for individuals that dispersed early versus late, but found nothing revealing in these comparisons (unpubl. data).
Dispersal Distance and Annual Reproduction
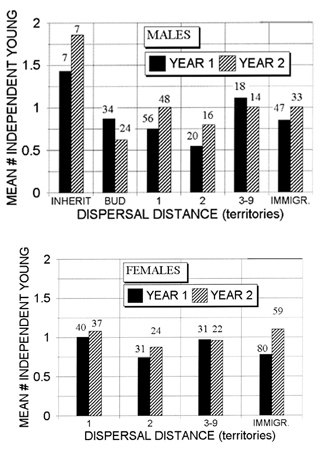
Dispersal distance did not appear to affect subsequent reproductive success when measured on an annual basis. Reasoning that any such effect would most likely exist early in the breeding lifespan, we compared annual reproduction during the first two years of breeding among jays of different dispersal classes. Jays of both sexes produced about 0.9 independent young per year regardless of dispersal distance (Fig. 3), except that reproduction by the few males that inherited their natal territory averaged higher than the rest (1.64 vs. 0.85 independent young per year for years 1 and 2 pooled; t = 2.1, P < .03). Several other analyses involving additional breeding years after the first two yielded similar results.
Annual Survival and Dispersal Distance
For both sexes, annual survival declined with dispersal distance, but the pattern among males includes an unexpected feature. Among females that became breeders only 1 or 2 territories from their natal territory, average annual survival during the first two breeding years was 0.86 (117 survivors in 136 breeder-years). Among females dispersing 3 or more territories from home, including immigrants into our tract, annual survival during the same breeding years was only 0.76 (149 survivors in 196 breeder-years; Chi Square = 5.1, P < .025). Among males, excluding those that became breeders through territorial budding, comparable survival values were identical to those of females (136/159 = 0.86 for dispersers up to 2 territories from home, and 91/119 = 0.76 for longer-distance dispersers; Chi Square = 3.7, P = .05). Pooling the sexes (again excluding males that budded) yielded highly significant evidence for reduced survival among the longest-distance dispersers (Chi Square = 7.5, P < .005). Highest annual survival over the first two years of breeding was exhibited by the 7 males that inherited their natal territory outright (13 survivors in 14 breeder-years, or 0.93).
About 20% of males became breeders by establishing a new territory and pairing with a neighbouring or immigrant female, on space that the male’s natal group had defended during preceding years. This process, known as territorial budding (Woolfenden and Fitzpatrick 1978, 1984), results in extremely short natal dispersal and a male jay breeding on space that is at least partially familiar. Therefore, budding might be expected to confer annual survival comparable to that of males that inherit their natal territory outright. To our surprise, however, the reverse appeared to be true: budded dispersers fared poorly immediately following the establishment of their new territories. Annual survival over the first two years of breeding by this sample averaged only 0.72 (43 survivors in 60 breeder-years), which was significantly below the 0.86 annual survival rate for all other males dispersing within two territories of home (Chi Square = 5.6, P < .025). As a dispersal tactic, therefore, establishing a new territory by budding clearly carries elevated risk during the early breeding years, at least for certain individuals.
Survivorship of males that budded their territories increased considerably after the second year of breeding. Annual survival of budded males that survived to their third breeding year averaged 0.86 through year 10 (67 surviving out of 78 breeder years), compared with the above-mentioned 0.72 for budded males during their first two breeding years (Chi square = 4.2, P<.05). This very high survival rate during the later years by budded males contrasts with that of long-distance dispersers, which averaged only 0.76 annual survival between breeding years 3 and 10. Although this difference was not statistically significant (Chi Square = 2.61, P = .14), it would be sufficient to cause an enormous and biologically meaningful difference in the life table of jays. Indeed, high survivorship after the second breeding year resulted in a mean breeding lifespan for budded males that was not appreciably different from the other short-distance dispersal categories (4.48 versus 4.86 years, respectively; Table 2). Mean breeding lifespan of long-distance disperser males (3.58 years) was significantly shorter than that of short-distance dispersers including those that budded (t=1.75, P < .05).
Lifetime Reproduction and Dispersal DistanceAn ultimate measure of relative costs and benefits of alternate life history strategies is lifetime production of offspring. Lower survivorship among long-distance dispersers, if translated into significantly lower lifetime reproductive success, suggests active selection in favour of natal philopatry. Among our pool of known-age, known-first-breeding cohorts, both males and females do indeed provide such evidence (Table 3), although the pattern appears to be statistically significant only for males. Males that became breeders beyond 2 territories of their natal territory (including immigrants) averaged 30% lower lifetime reproductive success compared to those breeding within 2 territories of home (p < .04 for fledgling production, and P < .03 for yearling production). Among females, the difference was only about 20% lower among the longer-distance dispersers. Although the latter difference was not statistically significant ( < .08 for fledgling production, P < .14 for yearling production), the similarity between the trends of males and females suggests that the pattern is general among Florida Scrub-Jays of both sexes.
DISCUSSION
Florida Scrub-Jays exhibit natal philopatry as extreme as in any bird species known. One quarter of all breeding males inherit at least part of their natal territory, and more than 75% breed within a 2-territory radius of their birthplace. Over half of all breeding females also recruit within a 2-territory radius, although a few females do disperse many kilometres from home. We observe that the enormous bias toward short-distance dispersal in this species has a proximate explanation, namely, the distinctly home-based searching strategies of prebreeders. Most jays eventually become breeders by pairing with widowed jays on existing territories, yet prebreeding jays rarely depart permanently to search for these vacancies. Instead, they monitor the neighbourhood from within the natal territory and may engage in temporary forays out from the natal territory, returning regularly (Woolfenden and Fitzpatrick 1984, 1986). The process of successfully competing for a vacancy may take several years. Continued dependence on the natal territory as a familiar place of refuge during this searching period obviously limits the opportunity for successful dispersal beyond the immediate neighbourhood of the natal territory.
Habitat fragmentation does not appear to alter the basic dispersal strategy described above. Florida Scrub-Jays are extreme habitat specialists (Woolfenden and Fitzpatrick 1984, 1996; Breininger et al. 1995, 1996), and in many areas suitable habitat occurs in isolated patches, either because of natural patchiness of the underlying sandy soil or from secondary fragmentation imposed by humans (e.g. Stith et al. 1996). In these areas, dispersal often requires traversing unsuitable habitat, and therefore could lead to dispersal behaviour more typical of widely dispersing species, where prebreeders leave home permanently in search of vacancies. Indeed, Thaxton and Hingtgen (1996) showed that Florida Scrub-Jays in isolated suburban fragments tended to disperse farther (and leave home earlier) than did nearby jays occupying more continuous habitat. We, too, found that jays occupying a naturally fragmented landscape dispersed over greater distances than those in continuous habitat. However, the difference held only in terms of absolute distances across the ground. When dispersal distance was measured as occupied jay-territories traversed, the two dispersal curves were nearly identical. Our behavioural observations corroborate that even in these fragmented habitats, prebreeders either remain at home or return home often. Hence, just as in continuous habitat, breeding vacancies in the vicinity of home tend to be at least encountered, if not favoured, over more distant ones. We interpret this as evidence that the fundamental dispersal tactic of this species, namely, to monitor the immediate neighbourhood for vacancies while benefiting from the comparative safety and cached food supplies within the natal territory, is not altered by natural habitat fragmentation.
Who are the few long-distance dispersers? For a species with such a home-based dispersal strategy, one hypothesis is that individuals move progressively farther away only after attempting unsuccessfully to fill vacancies near home (e.g. Zack and Rabenold 1989). Our data fail to support this hypothesis. For both males and females, the longest-distance dispersers (those dispersing more than 9 territories from home) were significantly biased toward jays that left home permanently before age 2. A few of these even departed before age 1, a rare event in this species. We detected no features of family or neighbourhood composition that were correlated with these long-distance dispersers at the time of their permanent departure. We conclude that for a minority of jays -- mostly females -- the prebreeding period spent searching for vacancies within the natal neighbourhood is short or absent. These individuals choose instead to pursue an early dispersal strategy similar to that described for Red-cockaded Woodpeckers (Walters 1990, Walters et al. 1992a).
'Polymorphism' in dispersal tactics among individuals of the same population has been documented elsewhere in birds (e.g. Greenwood and Harvey 1982, Johnson and Gaines 1990, Haas 1995), including other co-operative breeders (e.g. Walters 1990, Walters et al. 1992a, 1992b, Noske 1991, Zack and Skutchberry 1992), and its genetic consequences can be considerable (Payne 1990). However, a convincing evolutionary explanation for this variation is still lacking. Early long-distance dispersers could be individuals living in low-quality territories or habitat patches (Stacey and Ligon 1987, 1991), or, conversely, they could be subordinate individuals that fail to win in competition for breeding vacancies amidst high-quality habitats (Zack and Rabenold 1989). Evidence from Florida Scrub-Jays supports neither of these hypotheses, as long-distance dispersers have departed from some of the most successful territories (as well as some of the least successful), and many of these long-distance dispersers depart well before attempting to win breeding space locally.
Our data on survivorship, reproduction, and lifetime output in relation to dispersal distance provide tantalising evidence for an evolutionary interpretation of philopatry in the Florida Scrub-Jay. Annual reproductive rates of jays of both sexes are not related to dispersal distance. In contrast, however, survival rates are significantly higher among those that breed closest to the natal territory. This relationship appears to translate into greater lifetime reproductive success among the shortest-distance dispersers, suggesting that long-distance dispersal is selectively disfavoured in this species. It is not yet clear whether dispersal distance has a heritable component in this species, and the data on this point from other bird species are equivocal (Greenwood et al. 1979, van Noordwijk 1984, Payne and Payne 1993). If a heritable component exists, then the survival costs for long-distance dispersal -- even after successful pairing on a territory -- would appear to favour short-distance dispersal rather strongly, providing an evolutionary explanation for philopatry in this species.
We believe these to be the first data for any bird species to suggest a significant effect (positive or negative) of natal dispersal distance on lifetime fitness. The question is extremely difficult to study in most species because the longest-distance dispersers are difficult to locate, let alone be tracked throughout their lives (Newton 1989, Johnson and Gaines 1990). Comparing Red-cockaded Woodpeckers that remained home to breed versus those that dispersed, Walters et al. (1992a, 1992b) found no differences in annual or expected lifetime success. Studies of reproductive success following breeding dispersal (i.e. changes in breeding location from one breeding year to the next) suggest that individuals of many bird species may increase their annual and lifetime success by moving after nest failure (e.g. Beletsky and Orians 1987, Part and Gustofsson 1989, Grant and Grant 1992, Payne and Payne 1993). However, in none of these studies was the reproductive outcome compared among different classes of natal dispersal distance.
Data presented here do not establish a cause-effect relationship between dispersal distance and survival or lifetime reproduction. Specifically, we cannot rule out the possibility that both dispersal distance and adult survival rate are covariates of some other factor, or combination of factors, reflecting overall quality, site of origin, or other genetic or phenotypic feature of individuals. We do suspect, however, that the effect is real. If so, it is most likely caused by differential familiarity with both the physical environment of the breeding territory and the social environment surrounding the territory. Both of these factors can influence predation rate, and predation is the most important source of adult mortality in Florida Scrub-Jays. For a non-migratory species occupying year round territories, prior knowledge of such physical features as sentinel perches, spatial distribution of pine trees, productive foraging areas, and areas of optimal cover increases an individual’s ability to detect or escape a predator. Moreover, individuals are especially vulnerable to undetected attack by a predator during territorial display. Although not yet quantified, we have an impression that such displays are engaged in more frequently by jays that are relatively new to an area, and by jays that are attempting to establish new territorial boundaries. These features tend to characterise two classes of breeding jays: (1) long-distance dispersers, and (2) males in the process of budding and growing a new territory. These are the two classes exhibiting the lowest survival rates as breeders.
Survival rates among males that became breeders via territorial budding rose conspicuously after the second breeding year, a reversal that was not present in any other class of breeder, male or female. We suspect that this shift occurs through a combination of two related factors. First, competition for breeding space in this species is extremely intense (Fitzpatrick and Woolfenden 1986), hence most budded territories are small during their first year (Woolfenden and Fitzpatrick 1984). New territories require constant vigilance and frequent defence in order to persist and grow. Indeed, most territories that fail to grow over the first year or two disappeared by year three (Fitzpatrick et al. 1991). The process of maintaining and expanding new territories is energetically costly, and also exposes the breeders to elevated risk of surprise attack by a predator. Furthermore, new breeding pairs rarely are accompanied by helpers in the effort, because reproductive success increases with time spent together as a pair (Marzluff et al. 1996). The net cost of extra effort and exposure early in the existence of a territory is reflected in higher annual death rates among the breeders early after the budding process. We speculate that this difficult process of territory establishment exposes budded males to a form of 'culling,' in which lower-quality males tend to be eliminated during the difficult early years following a territorial bud, leaving higher-quality males over represented within the sample of later breeding years. Once they survive the first two years, these remaining males demonstrate survival rates comparable to or higher than all the other males breeding within two territories of their birthplace.
The dispersal strategies and outcomes described here are consistent with our long-standing contention that social behaviour in the Florida Scrub-Jay is driven by intense competition for breeding space in an environment where optimal habitat is abruptly bordered by habitat that is poor or unusable. The dispersal event itself is known to elevate mortality of prebreeders (Woolfenden and Fitzpatrick 1984, Fitzpatrick and Woolfenden 1986), and we have evidence that jays in fragmented habitat experience higher mortality during the dispersal period than those in continuous habitat (unpubl. data). Moreover, habitat patches beyond about 12 km from a source population are rarely colonised (Stith et al. 1996). These findings make it clear that dispersal in fragmented landscapes carries even greater cost than in continuous habitat. The natal territory serves as a vital refuge during this period, where sentinel and roosting sites are familiar and an abundant supply of cached acorns exists for each individual throughout the year. Data presented in this paper suggest that even after the dispersal event itself, living and breeding within the natal neighbourhood continues to confer survival advantages. Space is a crucial resource to Florida Scrub-Jays, and familiar space appears to be the most valuable of all. For all but the handful of jays that disperse inexplicably early, familiar space appears to be a resource for which jays are willing to spend up to several years competing.
REFERENCES
Beletsky, L.D. & Orians, G.H. 1987. Territoriality among Red-winged Blackbirds: I. Site fidelity and movement patterns. Behav. Ecol. Sociobiol. 20: 21-34.
Breininger, D.R., Larson, V.L., Duncan, B.W., Smith, R.B., Oddy, D.M. & Goodchild, M.F. 1995. Landscape patterns in Florida Scrub Jay habitat use and demographic success. Conservation Biology 9: 1442-1453.
Breininger, D.R., Larson, V.L., Oddy, D.M., Smith, R.B. & Barkaszi, M.J. 1996. Florida Scrub-Jay demography in different landscapes. Auk 113: 617-625.
Christman, S.P. & Judd, W.S. 1990. Notes on plants endemic to Florida scrub. Florida Scientist 53: 52-73.
Fitzpatrick, J.W., B. Pranty, and B. Stith. MS. Statewide distribution of the Florida Scrub Jay, 1993. Final report to U.S. Fish and Wildlife Service, Cooperative Agreement #14-16-0004-91-950.
Fitzpatrick, J.W., Woolfenden, G.E. & Kopeny, M.T. 1991. Ecology and development-related habitat requirements of the Florida Scrub Jay (Aphelocoma coerulescens coerulescens). v + 49 pp. Florida Game and Fresh Water Fish Commission, Nongame Wildlife Program Technical Report No. 8.
Fitzpatrick, J.W., & Woolfenden, G.E. 1986. Demographic routes to cooperative breeding in some New World jays. In: Nitecki, M., & Kitchell, J. (eds) Evolution of Behaviour. Univ. of Chicago Press, Chicago: 137-160
Grant, P.R. & Grant, B.R. 1992. Demography and the genetically effective sizes of two populations of Darwin’s finches. Ecology 73: 766-784.
Greenwood, P.J. 1980. Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 28: 1140-1162.
Greenwood, P.J. & Harvey, P.H. 1982. The natal and breeding dispersal of birds. Ann. Rev. Ecol. Sys. 13:1-21.
Greenwood, P.J., Harvey, P.H. & Perrins, C.M. 1978. Inbreeding and dispersal in the Great Tit. Nature 271:52-54.
Greenwood, P.J., Harvey, P.H. & Perrins, C.M. 1979. The role of dispersal in the great tit (Parus major): the causes, consequences and heritability of natal dispersal. J. Anim. Ecol. 48:123-142.
Haas, C.A. 1995. Dispersal and use of corridors by birds in wooded patches on an agricultural landscape. Conservation Biology 9:845-854.
Johnson, M.L. & Gaines, M.S. 1990. Evolution of dispersal: theoretical models and empirical tests using birds and mammals. Ann. Rev. Ecol. Syst. 21: 449-480.
Koenig, W.D., Pitelka, F.A., Carmen, W.J., Mumme, R.L. & Stanback, M.T. 1992. The evolution of delayed dispersal in cooperative breeders. Quart. Rev. Biol. 67: 111-150.
Marzluff, J.M., Woolfenden, G.E. , Fitzpatrick, J.W. & Balda, R.M. 1996. Breeding partnerships of two New World jays. In: Black, J.M. (ed.) Partnerships in birds. Oxford Univ. Press, Oxford: 138-16.
Mumme, R.L. 1992. Do helpers increase reproductive success? An experimental analysis in the Florida Scrub Jay. Behav. Ecol. & Sociobiol. 31: 319-328.
Newton, I. 1989. Lifetime Reproduction in Birds. Academic Press, London.
Noske, R.A. 1991. A demographic comparison of cooperatively breeding and non-cooperative treecreepers (Climacteridae). Emu 91: 73-86.
Pärt, T. & Gustofsson, L. 1989. Breeding dispersal in the Collared Flycatcher (Ficedula albicollis): possible causes and reproductive consequences. J. Anim. Ecol. 58: 305-320.
Payne, R.B. 1990. Natal dispersal, area effects, and effective population size. J. Field Ornith. 61: 396-403.
Payne, R.B. &. Payn, L.L. 1993. Breeding dispersal in Indigo Buntings: circumstances and consequences for breeding success and population structure. Condor 95: 1-24.
Stacey, P.B. & Ligon, J.D. 1987. Territory quality and dispersal options in the Acorn Woodpecker, and a challenge to the habitat-saturation model of cooperative breeding. Am. Nat. 130: 654-676.
Stacey, P.B. & Ligon, J.D. 1991. The benefits-of-philopatry hypothesis for the evolution of cooperative breeding: variation in territory quality and group size effects. Am. Nat. 137: 831-846.
Stith, B.M., Fitzpatrick, J.W., Woolfenden, G.E. & Pranty, B. 1996. Classification and conservation of metapopulations: a case study of the Florida Scrub-Jay. Pp. 187-215 in McCullough, D.R. (ed.), Metapopulations and Wildlife Conservation. Island Press, Washington.
Thaxton, J.E. & Hingtgen, T.M. 1996. Effects of suburbanization and habitat fragmentation on Florida Scrub-Jay dispersal. Florida Field Nat. 24: 25-37.
van Noordwijk, A.J. 1984. Problems in the analysis of dispersal and a critique on its ‘heritability’ in the Great Tit. J. Anim. Ecol. 53: 533-544.
Walters, J.R. 1990. Red-cockaded Woodpeckers: a ‘primitive’ cooperative breeder. In: Stacey, P.B. & Koenig, W.D. (eds) Cooperative Breeding In Birds: Long term studies of ecology and behaviour. Cambridge Univ. Press, Cambridge: 69-101
Walters, J.R., Doerr, P.D. & Carter, J.H. III. 1992a. Delayed dispersal and reproduction as a life-history tactic in cooperative breeders: fitness calculations from Red-cockaded Woodpeckers. Am Nat. 139: 623-643.
Walters, J.R., Copeyon, C.K. & Carter, J.H. III. 1992b. Test of the ecological basis of cooperative breeding in Red-cockaded Woodpeckers. Auk 109: 90-97.
Woolfenden, G.E. & Fitzpatrick, J.W. 1978. The inheritance of territory in group-breeding birds. Bioscience 28: 104-108.
Woolfenden, G.E. & Fitzpatrick, J.W. 1984. The Florida Scrub Jay: Demography of a Cooperative-breeding Bird. Monogr. Pop. Biol. No. 20, Princeton University Press, Princeton. 406 pp.
Woolfenden, G.E. & Fitzpatrick, J.W. 1996. Florida Scrub-Jay (Aphelocoma coerulescens). In: Poole, A. & Gill, F. (eds) The birds of North America, No. 228. Acad. Nat. Sci., Philadelphia and Amer. Ornithol. Union, Wash., DC: 1-28
Woolfenden, G.E. & Fitzpatrick, J.W. 1986. Sexual asymmetries in the life history of the Florida Scrub Jay. In: Rubenstein, D. & Wrangham, R.W. (eds) Ecological Aspects of Social Evolution: Birds and Mammals. Princeton Univ. Press, Princeton: 87-107.
Woolfenden, G.E. & Fitzpatrick, J.W. 1978a. The inheritance of territory in group-breeding birds. BioScience 28: 104-108.
Zack, S. & Rabenold, K.N. 1989. Assessment, age and proximity in dispersal contests among cooperative wrens: field experiments. Anim. Behav. 38: 235-247.
Zack, S. & Skutchberry, B.J. 1992. Delayed breeding in avian social systems: the role of territory quality and 'floater' tactics. Behaviour 123: 194-219.
Table 1. Dispersal distance and dispersal age in the Florida Scrub-Jay
Table 2. Breeding lifespans (yr) of Florida Scrub-Jays with different dispersal distances
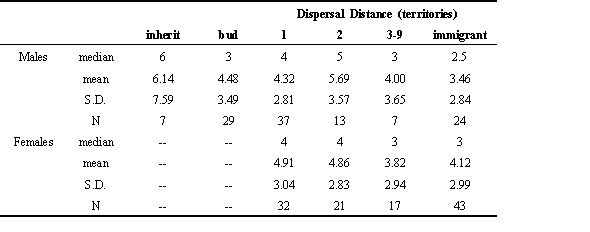
Table 3. Lifetime production of independent young in relation to dispersal distance

Fig. 1. Natal dispersal distances of male and female Florida Scrub-Jays color-ringed as nestlings or juveniles at Archbold Biological Station, 1969 - 1994. Distances represent actual territories traversed between natal and breeding territories. Dispersal distance of zero refers to individuals that inherited their territory; the next bar refers to those that budded (dispersal distance = 0.5 territories).
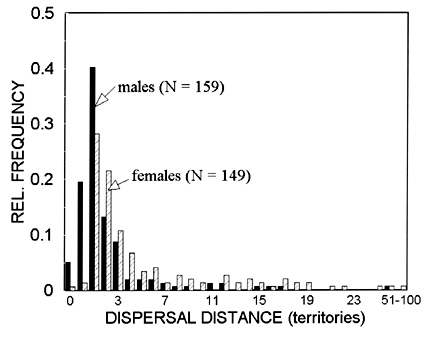
Fig. 2. Natal dispersal distances of Florida Scrub-Jays (sexes pooled) in continuous vs. fragmented habitat, where dispersals were measured in linear distance (Fig. 2a) and in territories traversed (Fig. 2b). The latter measurement produced statistically similar distributions.
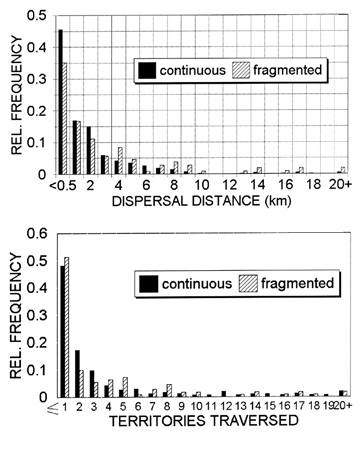
Fig. 3. Mean annual production of independent young by male (Fig. 3a) and female (Fig. 3b) Florida Scrub-Jays during their first and second breeding years following natal dispersals of different distances. Most immigrants to the tract ('immigr.') dispersed more than three territories. Sample sizes are shown above respective columns.