S40.2: Rigid and flexible adjustments to a periodic environment: Role of circadian and circannual programs
Eberhard Gwinner
Forschungsstelle für Ornithologie der Max-Planck-Gesellschaft, von der Tannstr. 3-7, D-82346 Andechs, Germany, fax 08152 373 33, e-mail gwinner@erl.ornithol.mpg.de
Gwinner, E. 1999. Rigid and flexible adjustments to a periodic environment: Role of circadian and circannual programs. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 2366-2378. Johannesburg: BirdLife South Africa.Endogenous daily (circadian) and annual (circannual) rhythms are ubiquitous mechanisms used by organisms to adjust physiological and behavioural functions to a periodically changing environment. Circadian and circannual adaptations are usually 'hard - wired' in the sense that both the basic oscillatory processes and the overt functions they control can hardly be modified by environmental conditions. However, in other cases these biological rhythms merely provide temporal frameworks within which the organism is free to adjust its performance according to circumstances. Thus by making use of their circadian clock, Sparrows can learn the time of day during which food becomes available. Circannual rhythms provide 'seasonal windows' within which individual variability is possible: the window for reproduction of the East-African Stonechat defines onset and end of the annual breeding season but within this window the bird can decide whether to breed once or twice. Likewise, the Garden Warbler's circannual program for migration determines when to begin and end migration; at the same time it permits the bird to respond flexibly within this scheme to acute environmental conditions and to modify its performance by learning. In thus allowing adaptation to a periodic environment, the fixed and variable elements of these biological clocks reflect the predictable and variable components of the environment in which the mechanisms have evolved.
INTRODUCTION
Birds, like most other organisms, are exposed to conspicuous diurnal and annual environmental changes and, as a consequence, have adjusted their physiology and behaviour to the periodic variations of those external factors that influence their fitness. Adaptations to the daily cycle are, for instance, reflected in the day-night alternation between wakefulness and sleep, and in numerous accompanying processes such as body temperature, food intake, or brain function. Adaptations to the environmental year manifest themselves, e.g., in the existence in most birds of particular breeding and moulting seasons, or in the annual recurrence of migrations and the host of morphological, physiological, and behavioural changes that are required for these journeys. While the close match between these biological rhythms and periodic changes in environmental conditions was initially taken as evidence for a direct and exclusive control of biological rhythms by environmental cues, it was recognised early on and generally accepted in the late ´50s and early ´60s that many daily and annual rhythms are not purely exogenously driven but rather have an endogenous, circadian or circannual, basis. Circadian rhythms have been found to control literally all 24-h biological cycles so far investigated, ranging from basic metabolic functions to complex behaviour (e.g., Aschoff 1981). Circannual rhythms are less widespread and necessarily restricted to the small percentage of organisms that live for one year or longer; still, circannual rhythms have now been reported for a considerable number of organisms including many birds, where they control functions as diverse as reproduction, moult, migration, body weight, and a multitude of less obvious parameters like the amplitude of the 24-h melatonin rhythm (e.g., Gwinner 1986).
With the discovery of endogenous 'circa-rhythms' as the basis of overt daily and annual rhythms in physiology and behaviour, attention was increasingly focused on the analysis of the mechanisms, both physiological and molecular, that generate these rhythms. In the circadian field organs and/or tissues have been found that contain pacemakers with a period of about 24 h, and genes have been identified that are essential for the production of rhythmicity at the cellular level. Although environmental factors have been identified that synchronise circa-rhythms with the periodic environment, other kinds of interactions between endogenous mechanisms and environmental stimuli have been largely neglected. Such interactions are, however, widespread and of paramount biological significance. Indeed, it turns out to be a major task of circa-rhythms to channel environmental stimuli into periodically changing, biologically meaningful responses. Endogenous and exogenous rhythms represent functional units. Therefore, to understand adaptations to rhythmic environments, the interactions between them must be thoroughly investigated. In some instances, these interactions provide the basis for immediate reactions: they determine, in a phase-dependent manner, the intensity of the response to an environmental stimulus; in extreme cases in which a particular response can only be elicited within a certain range of phases, the rhythm is said to provide a circadian or circannual 'window'. In other instances circa-rhythms enable the memorisation of periodically recurring, biologically meaningful events, thus allowing animals to perform appropriately during subsequent cycles. Such learned temporal patterns may be of a transitory nature or they may be retained for a long time, even for life.
Circannual programs: periodically changing dispositions to respond to environmental cues.
Circannual rhythms have so far been mainly studied in two groups of birds: tropical species and temperate-zone long-distance migrants (Gwinner 1986, 1996a, b). Among the former, emphasis has been laid on the circannual control of reproduction and moult; among the latter research has focused on the involvement of circannual components in the regulation of migratory activity. Both sets of studies exemplify how circannual rhythms help birds adjust to seasonal environments by simultaneously providing fixed elements and enabling variable responses.
a) Stonechats: circannual time programs for reproduction and moult.
Like their conspecifics in Europe, tropical Stonechats of East Africa Saxicola torquata axillaris are seasonal breeders. The testes of first-year Kenyan birds develop during the main dry season, October through March; as a consequence, birds are ready to start reproduction at the onset of the short rains, in March or April. Usually Kenyan Stonechats breed only once. Following the reproductive season gonads regress and birds initiate postnuptial moult. Two to three months after its completion gonadal growth is reinitiated (Dittami and Gwinner 1985).
The annual rhythms of gonadal size and moult have been found to persist for up to ten years even in birds kept in a constant 12.25 h equatorial photoperiod and under rigorously controlled temperature and dietary conditions (Gwinner and Dittami 1990; Gwinner 1996a). The period then deviates from 12 months, attesting to the endogenous nature of the underlying rhythmicity. Normal rhythms develop even in individuals exposed from early embryonic life to a constant laboratory environment and raised by parents which themselves had been kept in constant conditions for one year prior to the birth of their young (Gwinner 1995). This indicates that circannual rhythms are not only endogenous but also innate in the classical sense.
Circannual rhythms determine the time course of seasonal events in a genetically fixed subspecies-specific manner. A well-investigated example is provided by the temporal pattern of postjuvenile moult. If kept under the photoperiodic conditions of their respective breeding grounds, East African birds take longer to moult than their European conspecifics (S. t. rubicola). This difference is maintained if birds of the two subspecies are kept in the same constant environment. F1-hybrids show intermediary patterns (Gwinner and Neußer 1985).
In East African Stonechats, not only the moult but also the duration of the annual breeding season depends primarily on an endogenous circannual program. This is indicated by the observation that in a constant equatorial photoperiod the duration of the reproductively active phase, termed 'reproductive window', is similar to the reproductive window of freeliving conspecifics (Gwinner 1991). The reproductive window is defined as the interval between successive phases of gonadal growth and regression. Since in our captive birds nutritional conditions were constant and presumably much better than those of their conspecifics in nature, these observations suggest that favourable feeding conditions have no major effect on the duration of the width of the reproductive window.
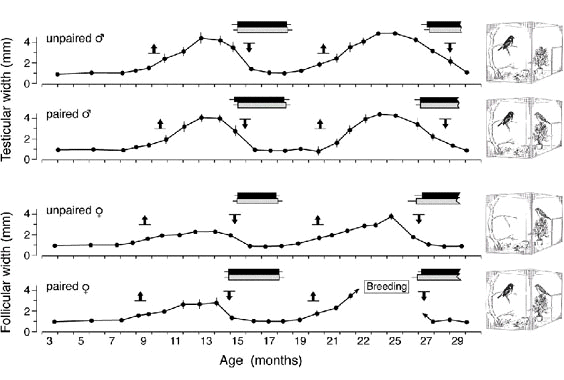
The width of the circannual reproductive window also turned out to be highly insensitive to social factors important for reproduction, particularly the presence and state of a mate. This is shown by the results of an experiment in which male and female African Stonechats were kept in individual aviaries, either singly or in pairs (Fig. 1). Photoperiod and other environmental factors were maintained constant over the 2 1/2 - year experimental period. All birds underwent two cycles of gonadal growth and regression, each of which was followed by a complete moult. However, while among the 10 individually-kept females only one built a nest and laid eggs, nine of the ten paired birds built nests and laid eggs, 48% of which were fertilised; from 44% of the eggs young hatched. Moreover, most pairs made more than one breeding attempt (Gwinner et al. 1995). As mentioned above, free-living Kenyan Stonechats of the population we have studied usually breed only once. In contrast, we frequently had captive pairs in this and another study (König and Gwinner 1995) that reproduced twice and, in one case, even three times per season. In addition, double-broodedness has been found to be common in another East African Stonechat population living in Tanzania only 250 km south of the Kenyan population (Scheuerlein unpublished observation). Taken together, all these observations suggest that within the framework provided by the circannual reproductive window, African Stonechats are rather flexible: depending on whether conditions are favourable or not, the birds reproduce or not; if they reproduce, they may raise two broods. This flexibility in the performance of the birds within the reproductive window is in sharp contrast with the rigidity in the width of the reproductive window itself: the gonads of the paired birds grew and regressed on average at the same time as those of the unpaired birds (Fig. 1). The favourable conditions for raising young did not result in a prolongation of the breeding season.
In a sense the combination of fixed and adjustable components of the circannual rhythm found in tropical Stonechats reflects the predictable and variable components of the seasonal environment to which stonechat circannual rhythms are adjusted. On a long-term basis, i.e., averaged over years, the equatorial breeding habitat of the Stonechat is highly predictable (as reflected, e.g., in the rainfall patterns published in Brown and Britton 1980), and it is likely that the circannual reproductive programs of equatorial Stonechats are adaptations to these long-term means of environmental patterns. Free-living Kenyan Stonechats have been shown to live for at least 10 years, and it would seem reasonable that lifetime reproductive success in such long-lived birds is enhanced by a strategy in which the putative breeding season remains in synchrony with the long-term average of the environmental cycles. The alternative strategy of shifting or extending the reproductive window according to the conditions prevailing in a particular year might be momentarily advantageous but disadvantageous in the long run: by extending the breeding season in one year (and as a consequence delaying ensuing post-reproductive events like moult) birds may partly or completely miss the breeding season of the subsequent year. - On the other hand, the high flexibility granted by the circannual clock within the framework of the reproductive window allows birds to either exploit favourable conditions by breeding twice, or to reduce, and perhaps even refrain from, reproductive activities when conditions are unfavourable (Fig. 2).
b. Warblers: time programs for migration.
Most Warbler species of the genera Phylloscopus and Sylvia are migratory, breeding in the temperate zones of Eurasia and wintering in more southerly regions, some travelling as far as South Africa. Migratory activity and accompanying processes like fat deposition as well as moulting and reproduction of these birds are controlled by a circannual rhythmicity (Gwinner 1968, 1986, 1996; Berthold 1988, 1993). The long- and medium-distance migrants travel more or less exclusively at night. If kept in cages, they exhibit nocturnal activity during the migratory seasons, and it has been known for some time that the intensity and duration of fall migratory activity is proportional to the distance normally covered on migration, particularly in autumn. These differences are even found if birds of the various species or populations are kept in the same constant environment (Gwinner 1968b; Berthold 1973). This fact, together with results of cross-breeding experiments revealing intermediary patterns in F1-hybrids between long- and short-distance migrants (Berthold and Querner 1981), indicates the existence of genetically determined species - or population-specific circannual programs for migration. Apart from specifying duration and overall amount of fall migratory restlessness, these programs also appear to control the temporal and perhaps even the spatial course of migration (Gwinner and Wiltschko 1978, 1980; Helbig et al. 1989); that is, seasonal changes in both migratory speed and migratory direction are under circannual control. In view of these and other findings it is likely that the temporal circannual programs play a major role in determining the distance and the time-course of migration in first-year migrants.
Like the reproductive schedules of Stonechats, the migratory time-programs of Warblers turn out to be rather rigid and resistant to some major manipulations. In Garden Warblers Sylvia borin, the prevention of migratory fat deposition at the beginning of the migratory season did not interfere with the temporally appropriate beginning of migratory restlessness (Berthold 1977). Correspondingly, when the body mass of Garden Warblers was reduced by starvation and kept low for several weeks, the end of fall migratory restlessness was not affected (Berthold 1976). Likewise, the end of migratory restlessness was not delayed in Garden Warblers whose nocturnal activity was suppressed either by keeping them in complete darkness during the night (Gwinner 1974) or by repeatedly withdrawing and returning food (Gwinner et al. 1992, see below).
While the temporal limits of the migratory timing programs are thus rigidly fixed, there are a few rather specific factors, or constellations of factors, that can modify the intensity of nocturnal activity within the framework of the circannual migratory program. One of these factors is food availability and/or the resulting increase or decrease in body mass (Biebach 1985). Figure 3 shows that Garden Warblers deprived of food during the fall migratory season exhibit a transitory increase of nocturnal migratory restlessness concomitant with a decline in body mass. During the subsequent phases of refeeding, migratory restlessness is interrupted. In the field, these responses presumably help migrants cope with changing nutritional conditions: intensifying actual migratory activity when food is scarce may increase the chance for these birds to find resting places favourable for refeeding; interrupting migratory activity when food becomes available following a phase of starvation may enable birds to replenish fat reserves depleted during the preceding phase of migration. Observations at stopover sites strongly support these suggestions (Biebach et al. 1986).
During the migratory season the increase in nocturnal activity following food withdrawal is not constant but depends on the phase of the cycle (Fig. 4): the increase is high early during the migratory season, declines towards the end of the migratory season and becomes essentially zero at the end of the season, i.e. around the time birds start moulting. Just as African Stonechats no longer respond to stimuli favourable for reproduction once the reproductive window has closed, Garden Warblers no longer respond to stimuli enhancing migratory activity once the migratory program has run down.
Circannual programs: modification and supplementation by learning.
Apart from providing the basis for specific phase-dependent responses to environmental cues, circannual rhythms are substrates for more permanent modifications of seasonal patterns that result from learning. Modifications of this kind are particularly well known from those migratory birds in which the endogenous timing programs that guide young birds on their first migrations are replaced or supplemented in older birds by mechanisms based on previous experience.
The well-known phenomenon of wintering-site fidelity provides striking examples. Even long-distance migrants like Garden Warblers, Reed Warblers Acrocephalus palustris, or Barn Swallows Hirundo rustica have been recaptured in winter at the site where they had been ringed during previous winters. Displacement experiments like those of Perdeck with European Starlings Sturnus vulgaris indicate that learning must be involved in the fixation of the wintering grounds. When caught and experimentally displaced perpendicular to their normal fall migratory direction, the adult birds compensated for this displacement, to migrate towards their normal wintering area by flying in an unusual direction. Young birds, in contrast, migrating for the first time in their lives, continued to fly in their original direction - to end up in a 'wrong' wintering site not normally visited by birds of this population (Perdeck 1964). Some of these birds were recovered in later years in the area in which they had spent the previous winter as a result of the displacement (Perdeck 1981). This supports the conclusion drawn from the initial result, that the birds had somehow learned the position of the site at which they had spent their first winter. The underlying navigational mechanism must be complex because the birds were obviously capable, upon release at an unfamiliar site, of determining their position relative to their goal area. It is as yet unclear to what extent these new mechanisms based on learning replace the original ones based on a timing program. Would adult birds, if transported to their wintering grounds soon after the beginning of fall migration, terminate migration instantaneously, having recognised the coordinates of their previous wintering grounds? Or would they continue to migrate, driven by their endogenous timing program?
Many cases of wintering-site fidelity have been reported from species like Sylvia or Acrocephalus Warblers that travel individually, unaccompanied by experienced adults (e.g., Curry-Lindahl 1981). In such species, learning of spatial properties of their wintering grounds must be exclusively based on interactions of the individual bird with its environment. The situation is obviously different in gregarious migrants, in which young birds follow experienced adults of a population or, in some species, their own parents. In such cases population - or family - traditions develop, as occurs for example in waders and ducks, or in geese and cranes. Traditions of this kind result in the return to specific wintering sites and even to particular resting areas along the migratory route. The learned nature of such species-specific migratory patterns becomes evident when certain stopover places are destroyed, or new ones are created. Under such conditions, novel routes develop within a short period of time (Berthold 1993; Alerstam 1990). So far, the mechanisms underlying such migratory route traditions have only rarely been studied. Recent attempts at imprinting goslings onto a light airplane that serves as surrogate parent, and using it to guide the imprinted young to a particular wintering site, were successful and may provide a promising tool to investigate questions about the nature of the mechanisms at work (Lishman 1996). It is possible that endogenous temporal components are a priori less significantly involved in controlling migrations of such gregariously migrating species. However, some kind of an annual clock must be involved in such species as well: learning of particular sites can serve the function of guiding birds on their migrations only if the learned place becomes associated with a particular time of the year.
Circadian time-place learning.
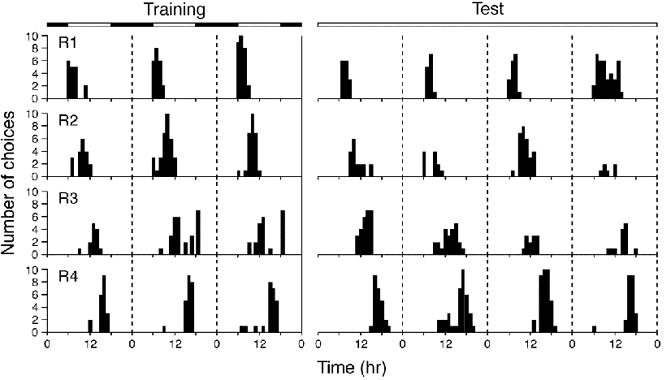
As early as 1910 Forel observed bees appearing on his garden table at breakfast time to feed on his marmalade. He was struck by the fact that the bees even appeared on the table at the appropriate time when no marmalade was available. These were the first suggestions of the existence of what von Frisch (1965) later called a 'time sense', the capacity to memorise the times of day at which food is available at certain places. Subsequent training experiments confirmed the existence of such a mechanism (e.g. Beling 1929) and further studies eventually proved that it is based on a circadian rhythmicity. A study by Stein (1951) revealed that birds, like bees, can be trained to associate certain feeding places with certain times of the day, and Wenger et al. (1991) finally demonstrated the circadian basis of this time-place learning mechanism in birds. Figure 5 shows the activity recording of a House Sparrow Passer domesticus that was initially kept under a light-dark cycle and trained to search for food in 4 different rooms at 4 different times of the day. When, during the subsequent test phase in continuous dim light, food was continuously available in all 4 rooms the learned pattern persisted for 4 days, to the end of the experiment. Under these constant conditions, the rhythm 'free-ran' with a circadian period slightly shorter than 24 h.
It is as yet unknown to what extent this capacity for time-place-learning, now documented for at least six different bird species, is utilised by free-living individuals to organise their foraging behaviour and other activities. Daily routines, obviously based on learning, have been described in several cases, for instance in Kestrels Falco tinnunculus that foraged along the same traplines day by day (Rijnsdorp et al. 1981). Enright (1975) gives examples from captive birds and suggests that circadian rhythms may generally act like 'a circa-24-h closed loop on a tape recorder' on which events occurring at certain times of the day can be recorded and recalled the following day or days. Enright suggests that these 'repeated circadian patterns of behaviour may represent the real ecological meaning, the primary adaptive significance of biological clocks for higher animals.' While the circadian oscillation as such is endogenous and has properties that can only barely be modified by environmental conditions, the rhythm provides an important basis for learning and retaining daily recurring features in the environment. As a consequence, an animal can re-use strategies successfully employed on the previous day.
CONCLUSIONS AND PERSPECTIVES
As outlined above, circannual and circadian rhythms provide the basis for a number of diverse adaptive responses to periodically changing environmental conditions. These responses are more or less fixed in some instances and highly flexible in others. Circannual programs provide rigid temporal frameworks, for instance for reproduction and migration; within the limits of these frameworks birds have the capacity to show a variety of different responses, dependent on environmental conditions. Circadian rhythms provide the basis for daily time-place learning and it is likely that a comparable circannual time-place learning exists as well. The latter possibility has not yet been rigorously investigated, but it may be that migratory birds associate spatial information acquired during their migratory flights with particular phases of an endogenous annual clock. This would help birds travel along the same migratory routes they used the previous year. Experiments along these lines seem highly promising.
While more and more details about the molecular and physiological bases of endogenous rhythms are being revealed, particularly of circadian rhythms, studies on their adaptive significance are becoming increasingly scarce. However, our proper biological interpretation of the functional architecture of these rhythms crucially depends on investigations addressing the question of what these rhythms are good for, i.e., in what respect they function as biological clocks.
REFERENCES
Alerstam, T. 1990. Bird migration. Cambridge; Cambridge University Press.
Aschoff, J. (ed.) 1981. Biological rhythms. Handbook of Behavioral Neurobiology. New York, London; Plenum.
Beling, I. 1929. Über das Zeitgedächtnis der Bienen. Z. vergl. Physiol. 9: 259-338.
Berthold, P. 1973. Relationships between migratory restlessness and migratory distance in six Sylvia species. Ibis 115: 594-599.
Berthold, P. 1976. Über den Einfluß der Fettdeposition auf die Zugunruhe bei der Gartengrasmücke (Sylvia borin). Vogelwarte 28: 263-266.
Berthold, P. 1977. Über die Entwicklung von Zugunruhe bei der Gartengrasmücke (Sylvia borin) bei verhinderter Fettdeposition. Vogelwarte 29: 113-116.
Berthold, P. 1988. The control of migration in European warblers. Proc. 19th Internat. Orn. Congr., Ottawa , 215-249.
Berthold, P. 1993. Bird migration. A general survey. Oxford, New York, Tokyo; Oxford University Press.
Berthold, P. & Querner, U. 1981. Genetic basis of migratory behavior in European warblers. Science 212: 77-79.
Biebach, H. 1985. Sahara stopover in migratory flycatchers: fat and food affect the time program. Experientia 41: 695-697.
Biebach, H. et al. 1986. Interaction of bodymass, fat, foraging, and stopover period in trans-sahara migrating passerine birds. Oecologia 69: 370-379.
Brown, L.H. & Britton, P.L. 1980. The breeding seasons of East African Birds. Nairobi; E.A.N.H.S.: 164pp.
Curry-Lindahl, K. 1981. Bird migration in Africa. London; Academic Press.
Dittami, J. & Gwinner, E. 1985. Annual cycles in the African stonechat Saxicola torquata axillaris and their relationship to environmental factors. J. Zool. Lond. A 207: 357-370.
Enright, J.T. 1975. The circadian tape recorder and its entrainment. New York; Intext Educat. Publ.
Forel, A. 1910. Das Sinnesleben der Insekten. München; E. Reinhard.
Frisch, K. v. 1965. Tanzsprache und Orientierung der Bienen. Berlin; Springer.
Gwinner, E. 1968a. Artspezifische Muster der Zugunruhe bei Laubsängern und ihre mögliche Bedeutung für die Beendigung des Zuges im Winterquartier. Z. Tierpsychol. 25: 843-853.
Gwinner, E. 1968b. Circannuale Periodik als Grundlage des jahreszeitlichen Funktionswandels bei Zugvögeln. Untersuchungen am Fitis (Phylloscopus trochilus) und am Waldlaubsänger (P. sibilatrix). J. Ornithologie 109: 70-95.
Gwinner, E. 1974. Endogenous temporal control of migratory restlessness in warblers. Naturwissenschaften 61: 405.
Gwinner, E. 1986. Circannual rhythms. Heidelberg; Springer.
Gwinner, E. 1991. Circannual rhythms in tropical and temperate-zone stonechats: A comparison of properties under constant conditions. Ökol. Vögel 13: 5-14.
Gwinner, E. 1995. Entwicklung circannualer Rhythmen tropischer Schwarzkehlchen (Saxicola torquata axillaris) nach Bebrütung und Aufzucht unter konstanten Bedingungen. J. Ornithologie 136: 79-82.
Gwinner, E. 1996a. Circannual clocks in avian reproduction and migration. Ibis 138: 47-63.
Gwinner, E. 1996b. Circadian and circannual programmes in avian migration. J. Exp. Biol. 199: 39-48.
Gwinner, E. & Dittami, J. 1990. Endogenous reproductive rhythms in a tropical bird. Science 249: 906-908.
Gwinner, E. & Neußer, V. 1985. Die Jugendmauser europäischer und afrikanischer Schwarzkehlchen (Saxicola torquata rubicola und axillaris) sowie von F1-Hybriden. J. Ornithologie 126: 219-220.
Gwinner, E. & Wiltschko, W. 1978. Endogenously controlled changes in migratory direction of the garden warbler, Sylvia borin. J. Comp. Physiology A 125: 267-273.
Gwinner, E. & Wiltschko, W. 1980. Circannual changes in migratory orientation of the garden warbler, Sylvia borin. Behav. Ecol. Sociobiol. 7: 73-78.
Gwinner, E. et al. 1988. Effects of food-deprivation on diurnal migratory restlessness and diurnal activity in the garden warbler Sylvia borin. Oecologia 77: 321-326.
Gwinner, E. et al. 1992. The migratory time program of the garden warbler: Is there compensation for interruptions? Ornis Scand. 23: 264-270.
Gwinner, E. et al. 1995. Endogenous gonadal, LH and molt rhythms in tropical stonechats: effect of pair bond on period, amplitude, and pattern of circannual cycles. J. comp. Physiol. A 177: 73-79.
Helbig, A.J. et al. 1989. Migratory orientation of blackcaps (Sylvia atricapilla): population-specific shifts of direction during the autumn. Ethology 82: 307-315.
König, S. & Gwinner, E. 1995. Frequency and timing of successive broods in captive African and European stonechats Saxicola torquata axillaris and S. t. rubicola. J. Avian Biol. 26: 247-254.
Lishman, W. 1996. Father Goose. New York; Crown Publishers.
Perdeck, A.C. 1964. An experiment on the ending of autumn migration in starlings. Ardea 52: 133-139.
Perdeck, A.C. 1981. Zugvögeln auf der Spur. Alte und neue Aufgaben der Vogelberingung. Umschau 81: 745-748.
Rijnsdorp, A. et al. 1981. Hunting in the kestrel, Falco tinnunculus, and the adaptive significance of daily habits. Oecologia 50: 391-406.
Stein, H. 1951. Untersuchungen über den Zeitsinn bei Vögeln. Z. vergl. Physiol. 33: 387 - 403.
Wenger, D. et al. 1991. Free-running circadian rhythm of a learned feeding pattern in starlings. Naturwissenschaften 78: 87-89.
Fig. 1. Changes in testicular width and in the diameter of the largest ovarian follicle in male and female East African Stonechats (Saxicola torquata axillaris) kept paired or unpaired in individual aviaries. flight feather moult; body moult. Means ± SEM are given.
¯: Mean (± SEM) onsets and ends of gonadal cycle (defined as the times at which testicular width reached a value of 1.5 mm and follicular diameter a value of 0.8 mm during phases of gonadal growth and regression). Each of the eight to ten individuals or pairs in the three experimental groups was kept in a 110 x 85 x 100 cm aviary and exposed from nestling age for 29 months to a constant 12.25 h photoperiod and a constant temperature of 20° ±3° C. Gonadal size was repeatedly measured by laparotomy (after Gwinner et al. 1995).
Fig. 2. Scheme of the proposed relationships between the endogenously programmed circannual window for reproduction, the environment, and the propensity of East African Stonechats to breed either once or twice per season. It is assumed that irrespective of conditions, the phase during which birds are potentially able to reproduce (curves) begins and ends at the same time each year. However, whether or not breeding is actually initiated depends on the environmental conditions prevailing in any given year. When conditions favourable for reproduction - like, for instance, rains and the accompanying increase in food supply - occur late, birds also breed late and, as a consequence, only once (upper diagram). When favourable conditions start early, birds breed early and subsequently have the opportunity to raise a second brood, provided conditions remain favourable (lower diagram).
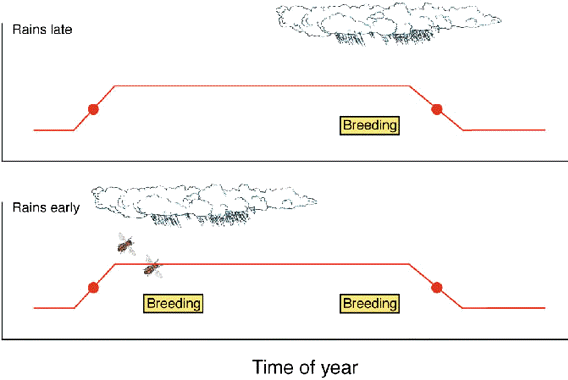
Fig. 3. Variations in nocturnal migratory restlessness (expressed as the number of 10-min intervals with activity per night; hatched areas) and body mass (curves) as well as the occurrence of winter moult (black bars) in 2 Garden Warblers (Sylvia borin) during the fall migratory season. Day 0 corresponds to September 1. The dotted segments of the curves show the body mass changes during food deprivation, the dashed segments those during the subsequent refeeding phase when the birds received 8 g of food per day, and the unbroken segments those during food ad libitum (after Gwinner et al. 1992).
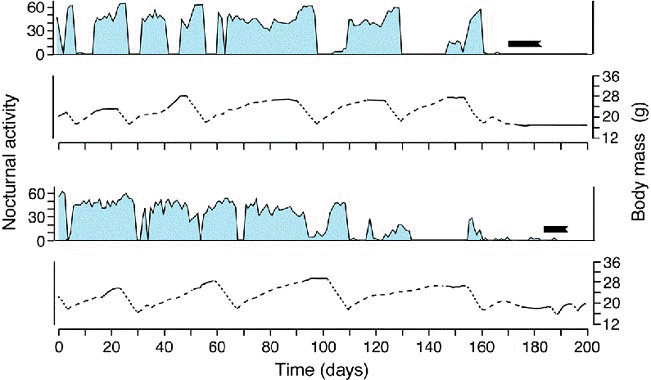
Fig. 4. Upper diagram: Variations of nocturnal migratory restlessness (expressed as the number of 10-min intervals with activity per night) of Garden Warblers. Horizontal bar: winter moult. Lower diagram: Changes in the duration of nocturnal migratory restlessness resulting from food withdrawal at different phases of fall migration. The bars show the increases in nocturnal activity during the first night following food withdrawal as compared with the average nightly activity exhibited during the last three nights preceding food withdrawal. In all cases medians with 95% confidence are shown (after Gwinner et al. 1988).
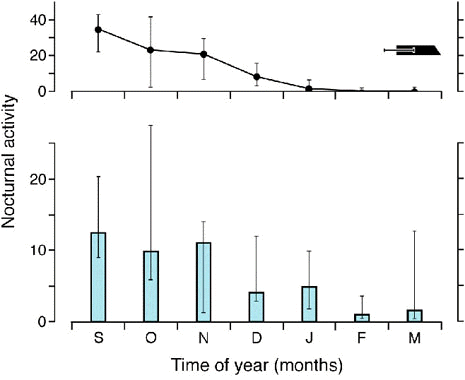
Fig. 5. Circadian time-place learning in a House Sparrow. The bird was kept in an apparatus consisting of a central living area connected to four feeding rooms (R1-R4). During the training phase (left), the bird was exposed to a 12:12 h light-dark cycle (lights 06:00 to 18:00), and food was available only in one room at a time in the following sequence: Room 1: 06:00 to 09:00; room 2: 09:00 to 12:00; room 3: 12:00 to 15:00; room 4: 15:00 to 18:00. During the test phase (right) the bird was exposed to constant dim light and food was continuously available in all 4 rooms. Each panel shows the number of times a particular room was chosen in each hour (Gwinner & Oltrogge unpublished).