S33.5: Bird migration across the Mediterranean
Bruno Bruderer & Felix Liechti
Swiss Ornithological Institute, CH-6204 Sempach, Switzerland, fax 41 41 462 97 10,
e-mail BrudererB@Orninst.ch; LiechtiF@Orninst.ch Bruderer, B. & Liechti, F. 1999. Bird migration across the Mediterranean. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1983-1999. Johannesburg: BirdLife South Africa.Migratory flyways between Europe and Africa are suggested to have evolved according to environmental conditions at different spatial and temporal scales: (1) at a continent-wide and long-term scale during the species’ range expansions following Pleistocene glaciation, (2) by the recent distribution of a) breeding and wintering ranges, b) ecological barriers and resting areas, c) the occurrence of predictable patterns of weather conditions, (3) by the evolution of behavioural reaction norms by which the individuals deal with the variability of conditions encountered en route. A trade-off between (A) risk-reduction by avoiding ecological barriers and (B) time/energy minimisation eventually leading to broad-front migration crossing barriers on the shortest routes. All this may have led to different solutions according to preconditions given by life-history traits. - Recent observations by radar, infrared, and large-scale coordinated moon-watching show important deviations of the migratory streams according to regional topography and suggest time-dependent risk-avoidance. Seaward migration decreases when the motivational and/or physiological state of the individual birds is due to deteriorate. - Tracking radar studies in southern Spain (near Malaga) and on the Balearic islands (Mallorca) reveal the diurnal pattern of nocturnal spring and autumn migration which is characterised by very short take-off phases. This allows inference about the take-off areas. In spite of probable concentration effects over the islands, the recorded density of passerine spring migration at the Baleares is only half of that at Malaga, wader/waterfowl migration only about one fifth; the latter difference emphasises the importance of the flyway along the African Atlantic coast.
INTRODUCTION
Bird migration between Europe and Africa is influenced by environmental conditions at different spatial and temporal scales. On a continent-wide and long-term scale, most bird populations were confined to the southern half of Europe during Pleistocene glaciations, the tree- and bush-dwelling species to the Mediterranean, their eastern and western populations at least partially separated by the tundra and steppe areas around the ice-covered Alps. This separation was responsible for the evolution of many sister species, races or distinct populations, which today often differ in their migratory directions. After the cold periods, the bird populations followed the progressing spread of vegetational zones associated with the climatic changes. Such range expansions required increasing migratory movements, the flyways developing according to the ways of range expansion. Many examples demonstrate that today’s migratory routes still reflect to a certain extent the ancient ways by which the birds had colonised their actual breeding ranges (Schüz 1971, Bruderer 1998).
At a medium spatial and temporal level, the flyways of landbirds are continuously moulded by evolution, the strongest selective pressures probably given by the distribution of topographical and ecological barriers, such as sea, desert, and mountain areas. Meteorological conditions may strongly influence the risk of mortality in critical areas. Such influence is most prominent in specialised birds such as soaring birds and birds depending on special habitats like wetlands. For birds which find suitable flight conditions and resting habitats all over Europe, one might expect that optimisation processes tended to minimise distance, energy or time for migration (Alerstam & Lindström 1990), time probably being less important in autumn than in spring. Current opinions, reflected in reviews and comprehensive books (Moreau 1972, Curry-Lindahl 1981, Lövei 1989), suggest that optimisation processes towards shorter routes resulted in a more or less homogeneous broad-front migration across the Mediterranean and the Sahara, particularly in nocturnal passerine trans-Saharan migrants. Support for this is derived from the pioneer surveillance radar study of Casement (1966), who found no evidence for concentrated nocturnal migration at any of the land bridges or narrows in the Mediterranean; however, reliable quantitative data are still lacking. Trapping data from small islands in the Mediterranean suggest that broad-front migration is a realistic assumption for some species (e.g. the Garden Warbler Sylvia borin) but not for others (Spina & Pilastro 1999).
A third level is given by the evolution of behavioural reaction norms which enable the individual birds to deal with the variability of environmental conditions en route. Many surveillance radar studies (Eastwood 1967) suggest that nocturnal migrants usually fly across coastlines without any deviation in their tracks. More detailed studies (Bruderer & Liechti 1998, Fortin et al. in press) showed, however, that there is considerable variation in the birds’ reaction to a coast depending particularly on the time of arrival in the coastal area.
In the present analysis we briefly review recent studies on the intensity and directions of nocturnal migration in the southern half of Europe. We then summarise available information and hypotheses in a map to provide a first synopsis of our ideas about nocturnal migration in relation to the three combined barriers of the Alps, the Mediterranean Sea, and the Sahara. In a second part of the study we will present quantitative data on the course of nocturnal migration in the western Mediterranean. We will, for the first time, compare the seasonal and diurnal variation in the intensity and composition of migration across the Mediterranean Sea and the Iberian Peninsula, respectively.
The main questions are: (1) from which areas and by what directions do nocturnal autumn migrants enter the area of the western Mediterranean? (2) do these directions indicate any adaptations to the distribution of the barriers of the Alps and the Mediterranean Sea? (3) do these directions suggest any behavioural reactions to topographical features? (4) what are, according to available information, the general directions of nocturnal bird migration in the southern half of Europe and what is suggested with respect to the distribution of the main passage areas? - In order to test the expected distribution of nocturnal migrants in two specific areas, we ask the following questions: (5) what proportions of nocturnal migrants cross the western Mediterranen Sea or fly across the Iberian Peninsula, respectively? (6) do these proportions shift within seasons, suggesting differences between trans-Saharan and pre-Saharan migrants?
METHODS
Our own studies are mainly based on three methods, calibrated against each other, and often used simultaneously: (1) coordinated, simultaneous moon-watching over large areas, (2) observations by a passive infrared camera, successively at various points along specifically defined transects, (3) observations by tracking radar at selected points.
A full description of the moon-watch method and its application in the area of the Alps was presented by (Liechti et al. 1996a). The method provides reliable data on migration traffic rates and directions of bird migration up to about 1000 m a.g.l.; it is, however, restricted to clear nights around full moon, and the opening angle of the field of view is only 0.5°. For the infrared observations we used a Long-Range-IR System (LORIS, IRTV-445L, Inframetrics, Massachusetts, U.S.A) aimed vertically upwards (Bruderer & Liechti 1994). This equipment provides information on migration traffic rates and directions up to 3000 m; the field of view is confined to an opening angle of 1.4°; flight altitudes are estimated according to silhouette size as in moon-watching; clouds reduce visibility. Simultaneous observations against the full moon by the pencil-beam of a tracking radar, by the infrared-equipment, and by a telescope provided a calibration of the observed silhouette sizes against the radar-measured distances (Liechti et al. 1995). A general review of radar as a tool for ornithological studies, with special emphasis on the radar used for the present studies, was given by (Bruderer 1997b). For more specific descriptions of the tracking radar 'Superfledermaus' (an ex-military fire-control radar with 3 cm wavelength, 150 kW peak pulse power, 2.2° nominal beam width, and 0.3-m s pulse length) with details on tracking and identification of single targets see (Bloch et al. 1981, Bruderer 1994); for details on the quantification of bird migration at various altitudes see (Bruderer et al. 1995).
Most of the second part of this paper is based on quantitative radar measurements (1) at the Andalusian coast 25 km east of Malaga (called Malaga site) and (2) close to the southern tip of Mallorca, the biggest of the Balearic Islands (called Baleares site). The measuring and analysis methods were similar to those described by (Bruderer et al. 1995). Generally, insect echoes can be excluded by a post-hoc application of a sensitivity time control STC to the recorded data (Bruderer et al. 1995). The very high density of insects at the Baleares site in autumn made it, however, impossible to determine migration density in the same way. Therefore, we restricted the analysis of autumn data at the present stage to some nights in late September and October with obviously low insect numbers. At both sites movements of local birds from or to the sea or along the coast were common during daytime, but on a reduced level also at night (e.g. swifts, gulls); reverse migration increased during the second half of the night (Bruderer & Liechti 1998). To calculate the intensity of forward migration we defined the proportion of 'netto migration' as the number of birds tracked, flying towards seasonally appropriate directions (within ± 90° of the mean migratory direction under calm winds) minus the number of birds flying towards other directions. This difference was divided by the total number of tracks. Thus, netto migration is the surplus of tracks with appropriate seasonal directions. This proportion differs strongly with height and time of day. For periods of ten days we calculated this proportion per each of five height intervals in diurnal time steps of two hours. Afterwards the echo densities for the corresponding height and time sequences were weighted by the proportion of netto migration. In the same way proportions of passerine- and wader-type migration were calculated. Quantities are, therefore, not directly comparable to those reported from central Europe (e.g. Bruderer & Liechti 1998) and southern Israel (Bruderer & Liechti 1995).
REVIEW OF RECENT RESULTS
Nocturnal autumn migration
Western Europe
The general appearance of nocturnal bird migration on the European mainland is a broad stream of migration roughly on a NE-SW axis, tending more SSW in areas closer to the Atlantic and more WSW along the coasts of the Mediterranean in northern Italy and southern France. To the W, the average directions may be influenced by western populations, e.g. those arriving from Britain on a southerly course as well as by birds from north-eastern populations shifting direction in western Europe. Migratory directions in northern Italy are influenced by the arrival of an important stream of birds from the southern part of eastern Europe (e.g. Hungary) and by a filtering effect of the Alps, leading to a considerable reduction of birds arriving from NE (summary by Bruderer 1997a). Due to the influx of migrants from upper Italy there is a considerable stream of migrants flying westwards along the coasts in southern France; this stream is complemented by strong migration following the northern border of the Alps and the Rhone Valley.
The area of the Alps
Results of 15 years of research in the area of the Alps, including southern Germany and northern Italy was reviewed by (Bruderer 1996): Due to basic directions of nocturnal autumn migration around SW (on average 230°) in central Europe, only a small part of this broad-front stream is far enough S to approach the Alps; this part slightly shifts direction according to the bow of the Alps and the Jura mountains. Important deviations from the basic direction are caused by the frequent westerly winds which drift the birds towards S and SE, and thus towards the Alps. Along the Alps the winds as well as bird migration are funnelled regionally by the mountain ranges and locally by single ridges. The influence of topography on the migrants increases with increasing headwind component, cloud cover and decreasing flight levels. High-flying birds have more southerly directions already above southern Germany. Birds above the Alps fly faster than those above the lowlands at the same altitude. According to ringing recoveries, birds arriving with southerly directions from far N are more often caught on Alpine passes than birds originating from central Europe. Birds caught in the Alps have more fat reserves and longer wings than their conspecifics in the lowlands. A coordinated moon-watch operation in a wide area N and S of the Alps, showed that even in good weather, with moderate opposing winds restricted to altitudes above 1500 m a.s.l., the concentration along the Alps was considerable, the density of migration to the S of the Alps reduced to 20% of that at the northern border of the mountain ranges. The directions S of the Alps were widely scattered, although mainly oriented towards W above northern Italy, shifting SW or S along the southward-bending ranges of the Mediterranean Alps (Liechti et al. 1996a, b).
The western Mediterranean
Basic information on the directions of migrants crossing the Mediterranean Sea was provided by (Casement 1966) using a ship-borne surveillance radar. He described a gap in migration above the Ionian Sea combined with NE/SW migration to the W of this gap and NNW/SSE directions to the E. (Rivera & Bruderer 1998) combined a recent series of infrared-observations along the Mediterranean coast from Toulon in southeastern France down to Tarifa in southwestern Spain with data from moon-watching (Wallraff & Kiepenheuer 1963), surveillance radar (e.g. Hilgerloh 1989, 1991), and tracking radar (Bruderer & Liechti 1998). These data indicate a general flow of migration on a NNE-SSW axis in the inland areas of France and Spain. Variation around this direction occurs in accordance with the actual wind situation. Scatter of directions and deviations increase when the migrants approach a mountain ridge. Along the Atlantic coast of southern France and Portugal, directions are more southerly in accordance with the general course of the coast. A comparison of the tracking radar and infrared data from the Balearic Islands and the Andalusian coast reveals prevailing southward directions on the islands and a temporal shift during night from SSW to W at the southern coast of Spain (Bruderer & Liechti 1998). An infrared transect along the European coast of the western Mediterranean (Rivera & Bruderer 1998) shows important variation of directions in accordance with the geographic situation. Southward or SSW-ward migration was observed along the southern coast of Spain (towards Africa) as well as to the south of the Massif Central and at the Costa Brava (towards the Balearic Islands and along the partially southward leading coast). Concentrated SW-migration followed the SE-coast of Spain.
Hypothetical flyways summarised
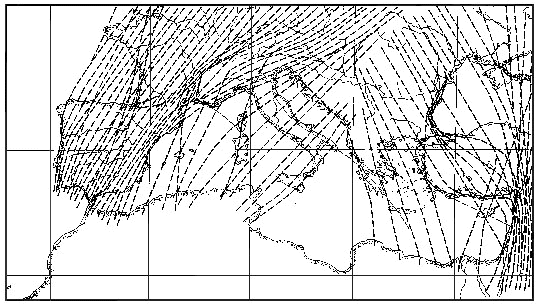
Fig. 1 summarises our ideas about the general streams of nocturnal migration in the southern half of Europe and the Middle East, using the punctual information provided by the publications mentioned above and some additional data from Israel (Alfiya 1995, Liechti & Bruderer 1995) as well as moon-watch data from North Africa (Kiepenheuer & Linsenmair 1965). Where no direct observations were available we derived interpolations from ringing recoveries (Zink 1973, Zink & Bairlein 1995). The map refers to autumn migration, which is more exposed to deviating effects of topography than spring migration. The information is simplified in many respects: It avoids crossing of lines representing SE-migration and the main stream towards SW, in spite of the well known fact that a few SE-migrants have breeding ranges reaching the Atlantic coast. Similarly, the lines crossing Italy on a NE-SW axis should start much further to the NE than indicated on the map. The lines can only suggest the principal directions in an area, each line symbolising the centre of a broad stream merging with the neighbouring streams. Concentrations of lines indicate the areas where deviations further upstream are supposed to lead to increased densities of migration.
The map suggests broad-front migration on a NE-SW axis across central Europe, with declining density to the SE. Directions are suggested to shift southward along the Atlantic coast, to be more westerly along the northern and southern borders of the Alps as well as along the French Mediterranean coast, and to shift back to SW or even SSW over the central parts of the Iberian Peninsula. Migration across the islands of the western Mediterranean as well as along the mainland of Italy cannot yet be quantitatively estimated, but is assumed to be non-negligible. The suggested concentration of migration along the eastern edge of the Mediterranean Sea is supposed to be mainly due to birds from areas to the N and NE of the Black and the Caspian Sea, many of them migrating on a S or SSW course. The map may be considered as a collection of hypotheses to be tested. The resulting streams of migration are suggested to be composed of a multitude of migratory streams of species and populations converging or diverging according to past or present environmental conditions; the whole assembly being best characterised by the term 'guided broad-front migration'.
Nocturnal spring migration
Data on spring migration are more scarce than for autumn. Hilgerloh‘s (1991) surveillance radar studies at Gibraltar show average directions very close to N, slightly shifted towards NNE in weak winds, and varying by about ± 20° around the still air directions under the influence of strong side-winds. Radar observations by (Hilgerloh 1987) and moon-watch data provided by Nisbet et al. (1961) indicate that spring migration is very weak over the Iberian Peninsula W of Cape Trafalgar; their catching data as well as moon-watch data suggest that most birds arriving in the western areas had been drifted by easterly winds. Houghton (1970) provides some evidence of local concentrations over the land areas at the Strait of Gibraltar. Nisbet et al. (1961) at Cape Trafalgar, recorded 32 to 80 birds in front of the moon during four sessions of two to three hours, resulting in migration traffic rates MTR of 700 to 2900 birds km-1 h-1; in one night they saw only 4 birds (MTR = 180); at Sanlucar, some 50 km further NW, they recorded practically no migration during similar observation periods.
Infrared observations of spring migration at the Andalusian coast showed on the one hand some W-E passage of birds following the coast and/or the mountain ranges of the Sierra Nevada, on the other hand important arrivals of birds from North Africa. During the arrival period of birds from North Africa average MTR was about 600, rarely reaching 1000 (unpubl. data); thus, roughly half of that observed at Cape Trafalgar. Observations near Hyères (east of Toulon) confirmed the autumn observations in the same area by showing important streams of migration arriving in southern France from the Costa Brava (on a WSW-NNE axis) and possibly from Sardinia (on a SE-NW axis), while only weak migration was noted approaching from the Balearic Islands (Rivera & Bruderer 1998); average MTR varied around 200, rarely reaching 500 to 800 birds (unpubl. data), thus about one third of that at the Andalusian coast. It seems that Toulon is already to the east of the stream of birds crossing the Balearic Islands, because near Aix en Provence Laty (1979) noted considerable arrivals from S, SSW and SW, those from Africa arriving only towards noon, therefore, having not been recorded by Rivera & Bruderer (1998). Spring migration on Mallorca was mainly towards N (Costa Brava), but with a notable proportion towards NE (Camargue area) according to a few nights with infrared observations in April 1995 (Bruderer et al. 1996). The same observations provided information on average migration traffic rates per night, which varied between 30 and 600, with an average of about 270 (Liechti et al. 1997), thus in a similar range as during early May in southern France, and somewhat less than half the MTR recorded at the Andalusian coast in mid-April.
New results
Moon-watch data from the western Mediterranean
Moon-watch data were collected in a coordinated operation during autumn 1996. The full-moon phase of September provided data from 42 sites, with 1749 single observations during a total of 145 hours (Fig. 2). Data from August and October were less complete, but did generally confirm the September data. They corroborate earlier observations in northern Italy (Liechti et al. 1996b) by showing wide directional scatter in the E of the Padanian Lowland (Site 1), westward migration over the main plain (Sites 2 and 4) and along the southern border of the Alps (3), shifting southward along the Mediterranean Alps (6), and following the coasts of the Italian and French Riviera (6, 7, 8); wide scatter of directions was recorded near Genova (6), Toulon (8) and at the Costa Brava (9). The intensities of migration observed in upper Italy were considerably higher than in previous observations; this indicates that temporally heavy passage from eastern countries occurs. Very weak SW-migration across the western Mediterranean basin is suggested by the observations near Rome, on Malta and on Mallorca. Directions in southwestern France (13) were mainly southward. The intensity of migration was lower in central Spain (14, 15) compared to southeastern Spain (16, 17, 18). Directions in southern Spain (16, 17) were rather SW than SSW, increased scatter occurring in the approach area to the Sierra Nevada (18). Average migration traffic rates in September varied around 1000 in upper Italy and southern France (Sites 1 to 8 and 13), around 1500 in southern Spain (Sites 16 to 18), and were below 500 near Rome (10) and on Malta (11).
Spring data were less complete than in autumn; therefore, we combined March, April, and May 1997, tolerating differences caused by season and by low numbers of observation nights at some sites (Fig. 3). The results include observations from 45 sites with 1193 single observations during 191 hours. A prevailing NNE tendency is recognisable in spite of wide directional scatter at most sites. Weak NE migration at the western edge of Sicily (4), near Rome (5), and near La Spezia (6) is caused by lack of arrivals across the sea during most of the observation time (2300 to 0100 hours), particularly at the Sites 4 and 5. Strong migration with prevailing NNE directions occurred in April at the Adriatic coast (7) and astonishingly strong migration was reported simultaneously from the centre of Italy (8). This high density is based on observations in three different valleys of the Apennin, where local topographical features concentrated migration and led it towards NNW and ESE/E. These three sites support the view that there is a notable stream of migrants flying along the Italian mainland. Near Nizza (9), ENE directions along the coast prevailed, while over the Venetian and Padanian plains (10 to14) directions were more NNE than expected according to autumn migration; wide scatter was particularly pronounced along the border of the Alps (15 to 17). Average migration traffic rates varied around 500, exceeding 1000 at some sites in upper Italy during March and 2500 (Site 8) in central Italy during April.
Quantitative radar data from Malaga and the Balearic Islands
Autumn migration
At the Malaga site 29866 individual flight paths were recorded and 124204 echoes from 1107 measurements were included to calculate migratory density from 11 August to 24 October 1996. Migratory activity was generally low in August. Wader/waterfowl migration reached levels similar to passerine migration in August and early September: it culminated in mid-September, and slowly declined in October (Fig. 4). Passerines reached a first peak simultaneously with the waders in September and declined in early October. However, a steep increase in passerine migration in mid- and late October suggested high numbers of pre-Saharan migrants such as Robin Erithacus rubecula, Black Redstart Phoenicurus ochruros Chiffchaff Phylloscopus collybita, larger thrushes Turdus ssp., Starling Sturnus vulgaris, and Skylark Alauda arvensis (the latter three represented in the radar data by a high proportion of large passerines).
The diurnal pattern of migration reflects the recruiting areas of the migrants. After an abrupt start of migration by birds having probably landed towards the end of the previous night in the coastal area and some dozens of kilometres inland from the Malaga coast (Fig. 5), migratory intensity declined steeply during the second and third hour after the start of migration. This decline was particularly pronounced during the migration of trans-Saharan migrants in September. In October the decline continued more steadily throughout the night. Migration ended about 11 hours after sunset in August and September, 13 hours after sunset at the end of October, corresponding to the shifting time of sunrise.
At the Baleares site there was nearly no bird migration in August, indicating very weak passage of trans-Saharan migrants. August and most September data were severely 'contaminated' by insects, which at this stage of the data analysis could not be 'cleaned'. Some nights in late September and in October (26 September to 3 October except the night of 28/29 September and 1/2 October; 14 to 24 October) were sufficiently clean to be used for a comparison of the diurnal migration pattern at Malaga and the Baleares. The curve from Malaga corresponds well with the October-curve of that site in Fig. 5. The curve from the Baleares is characterise by massive take-offs from the island. The subsequent decline is due to the gap in arrivals caused by the sea and by the subsequent arrivals of birds from different parts of southern Europe: About five hours after the first take-offs the influx from the Costa Dorada and southern parts of the Costa Brava (240 km to the NW) were recorded; culmination followed about seven hours after the start of migration by birds from the Costa Brava (300 km to the N); a second, smaller peak followed about eleven hours after take-off with birds from the Camargue area (some 480 km to the NNE). The volume of recorded migration at the Baleares was astonishingly high, i.e. much closer to the volume at Malaga than observed in April and May (see below).
Spring migration
From 22 March to 26 May 1997 30353 flight paths and 198947 echoes from 1539 measurements were included in the results for the Malaga site. At the Baleares site during the same time period 24085 flight paths and 296926 echoes from 4629 measurements were included (for part of the time density measurements were done parallel by two radars).
Passerine spring migration at Malaga (Fig. 6) showed a steady increase in density from late March to late April and a nearly symmetrical decrease throughout May. The density of migration was on average about twice that measured in the S of Mallorca (Baleares). Although most pre-Saharan migrants had already passed during February and early March the relative level of the sea-crossing migration was still high in late March and migration across the sea peaked slightly earlier in the season than landbound migration. Migration of birds with continuous wingbeats (mainly waders and waterfowl) was weaker than passerine migration, and almost five times stronger at the mainland compared to the island site.
The diurnal pattern of migration at coastal sites is determined by the distribution of land and water. The average course of bird movements throughout the night differs accordingly at the two sites (Fig. 7). Minimum intensity of northward movements was recorded at both sites just around sunset.
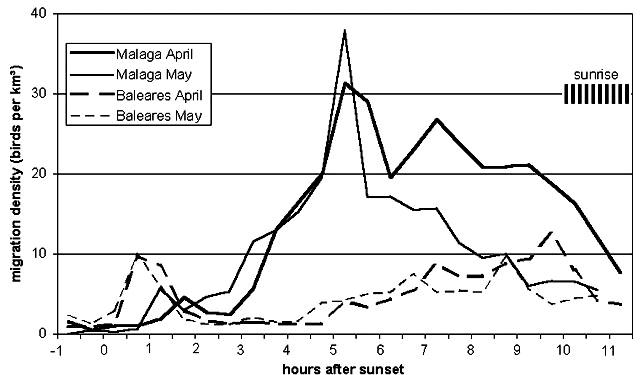
At the Malaga coast there was only weak migration along the coast during 2½ hours after sunset, until birds from North Africa arrived after flights of at least 170 km across the sea (Fig. 7). With wind support, the fastest and earliest departing birds arrived about three hours after sunset. Peak migration was reached 5 hours after sunset with birds having probably started between 30 and 60 minutes after sunset. This is confirmed by the average measured ground speeds of 40 to 45 km h-1, which would take them about 4 hours to cross the sea (Bruderer & Liechti 1998). The take-off area of birds arriving 4.5 to 5.5 hours after sunset can, therefore, be assumed to be within a narrow strip along the North African coast. After this peak, migratory intensity declined sharply, indicating a strong concentration of migrants along the North African coast. This peak was more pronounced in May than in April. The very short departure times observed after sunset (see departures at the Baleares: Liechti et al.1997 and this paper) suggest that birds arriving between 5 and 11 hours after sunset had been taking off at distances between the North African coast and about 300 km inland, i.e. in the Atlas mountains. Note that in April a second peak was observed about 7 hours after sunset suggesting a concentration of migrants 100 to 150 km inland. Generally migration intensity declined earlier in May than in April, which indicates that a high proportion of trans-Saharan migrants rest along the North African coast.
At the Baleares site (Fig. 7) there was a short burst of intense departures from the small land surface to the S of the radar surveyed area (about 65 km2 including the Cabrera islands situated roughly 20 km south of the radar). Obviously, the number of birds having landed on this small surface during the previous night and probably early morning hours was very high. Two hours after sunset (i.e. about 1.5 hours after the start of the main take-off) density returned to levels as low as those around sunset. Five hours after sunset densities started to increase slowly due to birds arriving either from the Spanish mainland (roughly 250 km to the SW; some may also be expected from Ibiza some 150 km to the SW. Higher levels were reached six to seven hours after sunset, corresponding to flight times of about six hours and flight distances of 300 km (from N-Africa) with relatively high ground speeds of 50 km h-1 (Liechti & Bruderer 1995). Peak migration occurred 8 to 10 hours after take-off, i.e. after flights of 300 to 500 km with the average observed ground speed of 40 to 50 km h-1. Half an hour before sunrise migratory activity declined, probably because birds still arriving across the sea were steeply landing in the coastal strip to the south of the radar (contributing to massive take-offs in the subsequent evenings).
Infrared transect to the N of Gibraltar
Infrared observations at 120-190 km NW, N and NE of Gibraltar confirmed general SW/SSW-migration at all five sites. These directions prevailed during all nights at the three northeastern sites, while at the more western sites intensities of migration were lower due to less favourable weather and directions were widely scattered due to some nights with reverse or SE-migration. SE-migration at the NW-site was particularly pronounced after nights with easterly winds. Only at the coastal site near Malaga migratory directions shifted from SSW in the first half of the night to W/WSW, following more or less the coast in the second half of the night (Trösch et al. in press). This shift in direction towards morning, following SSW departures towards Africa, was observed consistently by radar in the same area (Bruderer & Liechti 1998). The data suggest that temporal shifts in directions, either in the course of the night or after flights leading the birds too far W, result in a slight concentration of nocturnal migration towards the Strait of Gibraltar in addition to prevailing SW/SSW migration.
DISCUSSION AND CONCLUSIONS
Review
Migratory directions towards SW and SE combined with an obvious lack of southward migration in central and northern Europe are corroborated by data derived from bird ringing (Zink 1973-1985), by cage experiments (Helbig 1992), and by radar observations (Baumgartner & Bruderer 1985, Alerstam 1990). This paper suggests that these SW and SE deviations in the European-African migration system are a consequence of adaptations at various temporal and spatial scales to the combined ecological barriers of the Alps, the Mediterranean Sea, and the Sahara. This contrasts with bird migration in North America where there is no ecological barrier across the main flyways north of 30° latitude and migratory directions are generally southward, with a tendency towards SW according to the situation of breeding areas relative to the inter-American land bridge. Migratory intensities strongly depend on the regular passage of cold fronts and the subsequent favourable wind conditions (Richardson 1978, 1990), directions shifting with the wind to a high degree (Able 1974, 1980). Due to prevailing westerly winds, the SW-migrants in Europe are often confronted with opposing winds; they can not be as selective for favourable winds as the North American migrants and should try to avoid too much deviation by winds, i.e. they should be more prone to compensate or correct for winddrift than birds in North America. The hypothetical map presented in Fig. 1 suggests basic directions of migration in different regions, adapted to the large scale geographical conditions. It assumes broad-front migration of nocturnal migrants which may temporally be deviated by winds and regionally or locally by topographical features. Relevant concentrations of migration result from the convergence of the flyways used by different populations and additionally by deviations over long distances (guided broad-front migration).
For the western Mediterranean, Fig. 1 suggests that migration across the sea is relatively weak in autumn, the directions more southerly in the western part of the basin compared to southwesterly directions over the Thyrrenian Sea (approaching across Italy). Concentrations of SW-migration are proposed along the SE coast of Spain, fanning out again over southeastern Spain. A concentration of migrants is also assumed along the W coast of the Iberian Peninsula due to an influx of birds from NNE and southward bending directions of birds from NE.
New data
Autumn migration
The new moon-watch data are consistent with earlier findings that the general directions of nocturnal migration lead to a convergence of migratory streams over the Iberian Peninsula. This general funnelling of directions is increased by deviations observed in coastal areas, particularly in upper Italy and southern France.
The infrared transect to the N of Gibraltar corroborated the general SW/SSW flow of migration in southern Spain and confirmed that migration observed by radar near Malaga is representative for the general flow of migration in southern Spain. Additionally the infrared observations suggest that diurnal and weather-dependent shifts lead to a slight concentration of nocturnal migrants towards the Strait of Gibraltar.
Radar observations at Malaga indicate that most trans-Saharan migrants do not start their flight across the sea and the desert before September. This corresponds to observations from Israel (Bruderer & Liechti 1995). The relatively high numbers of resting migrants on Gibraltar in August (Finlayson 1992) are probably not indicating passage, but rather birds staying in the area for longer times. The visually observed passage of certain trans-Saharan species at Windmill Hill, Gibraltar, confirms the weak passage in August; notable numbers of the following species are observed only in September: e.g. Turtle Dove Streptopelia turtur, Bee-eater Merops apiaster, Nightingale Luscinia megarhynchos, Redstart Phoenicurus phoenicurus, Whinchat Saxicola rubetra, Garden Warbler Sylvia borin, Pied Flycatcher Ficedula hypoleuca); some August passage is reported for Willow and Bonelli’s Warbler Phylloscopus trochilus and bonelli (Finlayson 1992). Heavy migration observed by radar towards the end of October indicates important populations of pre-Saharan migrants wintering in the Maghreb area. This contrasts to the Negev in southern Israel, where the numbers of pre-Saharan migrants are very low (Bruderer & Liechti 1995), reflecting the fact that the semi-desert of the Negev is already to the S of the wintering areas of most pre-Saharan migrants.
The diurnal pattern of migration at Malaga in September is characterised by a sharp decrease in density after an impressive take-off phase. This implies a high concentration of trans-Saharan migrants close to the coast. This concentration is not only caused by premature landing of birds approaching the coastal area late at night, but additionally by birds returning from the sea towards morning (Bruderer & Liechti 1998) and possibly by birds waiting for optimal weather conditions. The decrease in October was less pronounced: by shifting along the coast and continuing migration along the coast, the pre-Saharan migrants in October remained airborne for a longer time, but accumulated along the coast to equal numbers as the trans-Saharan migrants in September. This difference between migration in September and October may be due to different behavioural reaction norms of trans-Saharan and pre-Saharan migrants when confronted with the Mediterranean Sea. Pre-Saharan migrants may be more flexible with respect to flight directions in response to topography. Trans-Saharan migrants may cling more strictly to their chosen direction, their diurnal schedule inducing either landing in coastal areas or returning from initiated sea-crossings before daybreak; when already far from the coast they may continue migration into the day, as indicated by the arrival of spring migrants from Africa towards noon in southern France (Laty 1979).
The diurnal pattern of late autumn migration at the Baleares site reflects the passage of migratory waves having departed from various parts of the European coasts. Interestingly, the volume of departures from the island was about 75% of that at the Malaga coast. This may be caused by an increased convergence of approaching pre-Saharan migrants diverting from previous course when the island comes within view. Pre-Saharan migrants may also show increased flexibility with respect to local shifts in direction (see above), thus being funnelled over the island towards its southern tip where the radar measurements took place. On the other hand we might think of an increased propensity of pre-Saharan migrants to cross the Mediterranean Sea compared to trans-Saharan migrants. If this paradox will prove to be true, a possible explanation may be that the Mediterranean Sea with about 500 km width may be overflown even with limited fat reserves, while crossing the Sahara is a bold venture requiring good wind conditions in addition to enormous fuel reserves. Following the land bridge allows refuelling until immediately before the desert crossing and offers improved possibilities to choose optimal weather conditions for the desert crossing.
Spring migration
Obviously we missed the main passage of pre-Saharan migrants during late February and the first two thirds of March (Finlayson 1992). The relatively high proportion of large passerines during peak migration suggests, however, that migration of pre- and trans-Saharan migrants overlaps more than in autumn. This idea is supported by Finlayson’s (1992) observations on grounded migrants at Gibraltar, where the overlap between the two groups seems to be more pronounced in April than in September.
Our radar data show that from late March to late May the density of migration across the Mediterranean basin between Algeria and France is at least two to three times lower than that across the Iberian Peninsula, thus corroborating earlier observations by infrared. The real difference is probably much larger, because it is reasonable to assume that the island attracts birds, particularly towards the end of the night. The attractiveness of the island is reflected by the high numbers of birds departing in the evening from a surface of only 65 km2 with appropriate habitats on about 30 km2. According to the migration traffic rate (MTR) measured at the radar site the density of resting migrants to the S of the radar must have been about 5 birds ha-1, in some nights up to 30 birds ha-1 of suitable habitat. The difference between the amount of passage at the Baleares compared to Malaga is more pronounced for birds with continuous wing-beats, such as waders and waterfowl, than in passerines. The reduced passage of waders far away from the Atlantic is on the one hand understandable, because most species depend on coastal and estuarian habitats outside the breeding period. On the other hand, the alternative idea that these powerful trans-Saharan migrants would not hesitate to fly across large inhospitable areas in order to minimise the distance between breeding and non-breeding areas is very attractive, but has in this case to be rejected.
Moon-watch data indicated important migration along the Italian Peninsula and across the Adriatic Sea in some spring nights. The question, whether these birds are coming via Sicily and southern Italy or straight across the sea from Tunisia, cannot be answered by the available data, because observations at the western tip of Sicily (Site 4) and at the coast near Rome (5) where made during times of the night when not much migration could be expected at these coastal sites. Accumulation of birds on the Italian mainland during the rest of the night is possible. According to trapping data, direct flights over the sea are a common phenomenon (Spina & Pilastro 1999), but quantitative comparisons are still lacking.
The diurnal pattern at Malaga shows a narrow peak around 5 hours after sunset in May, indicating a strong concentration of trans-Saharan migrants along the North African coast. This is the analogon of the impressive evening departures from the Malaga coast in September. In April the second peak about 7½ hours after sunset might indicate important resting areas in the highlands of the Atlas. Detailed analysis of daily patterns including wind support will inform more specifically about particular take-off areas in Algeria and Marocco.
ACKNOWLEDGEMENTS
We are grateful to all the colleagues engaged in the studies carried out in the area of the Alps and in the western Mediterranean. The Swiss National Science Foundation supported both projects; currently by grant number 31-432 42.95. The Silva-Casa and Wolfermann-Nägeli Foundations supported parts of the Mediterranean project. We are grateful to the Spanish authorities for support at the national, regional and local level. The Swiss Army provided the radars. Th. Steuri developed the recording equipment; he and D. Peter kept the radars going during the field work. Many students cooperated in the field and analysed parts of the data. V. Keller, M. Kestenholz, D. Peter, and L. Schifferli corrected parts of the present text.
REFERENCES
Able, K.P. 1974. Environmental influences on the orientation of free-flying nocturnal bird migrants. Anim. Behav. 22: 224-238.
Able, K.P. 1980. Mechanisms of orientation, navigation, and homing. In: Gauthreaux, S.A. (ed.) Animal migration, orientation, and navigation: 283-373. London, New York: Academic Press.
Adams, D.W.H. 1962. Radar observations of bird migration in Cyprus. Ibis 104: 133-146.
Alerstam, T. 1990. Bird Migration. Cambridge: Cambridge University Press.
Alerstam, T. & Lindström, A. 1990. Optimal bird migration: the relative importance of time, energy, and safety. In Gwinner, E. (ed.) Bird migration: 331-351. Berlin Heidelberg: Springer.
Alfiya, H. 1995. Surveillance radar data on nocturnal bird migration over Israel, 1989-1993. Isr. J. Zool. 41: 517-522.
Baumgartner, M. & Bruderer, B. 1985. Radarbeobachtungen über die Richtungen des nächtlichen Vogelzuges am nördlichen Alpenrand. Orn. Beob. 82: 207-230.
Bloch, R., Bruderer, B. & Steiner, P. 1981. Flugverhalten nächlich ziehender Vögel - Radardaten über den Zug verschiedener Vogeltypen auf einem Alpenpass. Die Vogelwarte 31: 119-149.
Bruderer, B. 1994. Radar studies on nocturnal bird migration in the Negev. Ostrich 65: 204-212.
Bruderer, B. 1996. Vogelzugforschung im Bereich der Alpen 1980-1995. Orn. Beob. 93: 119-130.
Bruderer, B. 1997a. Migratory directions of birds under the influence of wind and topography. In: Orientation & Navigation - birds, humans & other amimals, Vol. 27: 1-10. Oxford: Royal Institute of Navigation.
Bruderer, B. 1997b. The study of bird migration by radar. Part 1: The technical basis. Naturwissenschaften 84: 1-8.
Bruderer, B. 1998. Proc. Symposium Conservation of Migratory Species. Gland: .
Bruderer, B. & Liechti, F. 1994. Quantification of bird migration - different means compared. Bird Strike Comm. Europe 22: 243-254.
Bruderer, B. & Liechti, F. 1995. Variation in density and height distribution of nocturnal migration in the south of Israel. Isr. J. Zool.: 477-487.
Bruderer, B. & Liechti, F. 1998. Etude des migrations transméditerranéennes au moyen du radar. Directions de la migration nocturne en automne prés de Malaga et à Majorque. Nos Oiseaux 45, suppl. 1: 49-58.
Bruderer, B. & Liechti, F. 1998. Flight behaviour of nocturnally migrating birds in coastal areas - crossing or coasting. J. Avian. Biol. 29: 499-507.
Bruderer, B., Steuri, T. & Baumgartner, M. 1995. Short-range high-precision surveillance of nocturnal migration and tracking of single targets. Isr. J. Zool. 41: 207-220.
Bruderer, B., Liechti, F. & Steuri, T. 1996. Migrations aviennes à travers l'ouest méditerranéen - direction de vol au printemps au-dessus de Majorque. Alauda 64: 7-16.
Casement, M.B. 1966. Migration across the mediterranean observed by radar. Ibis 108: 461-491.
Curry-Lindahl, K. 1981. Bird Migration in Africa., Vol. 1, 2. London, New York: Academic Press.
Eastwood, E. 1967. Radar Ornithology. London: Methuen.
Finlayson, C. (ed.) 1992. Birds of the Strait of Gibraltar. London: Poyser.
Fortin, D., Liechti, F. & Bruderer, B. in press. Variation in the nocturnal flight behaviour of migratory birds along the norhtwest coast of the Mediterranean Sea. Ibis.
Helbig, A.J. 1992. Population differentiation of migratory directions in birds: comparison between ringing results and orientation behaviour of hand-raised migrants. Oecologia 90: 483-488.
Hilgerloh, G. 1987. Zeitlicher und räumlicher Verlauf des Singvogelzuges. Courier Forschungsinstitut Senkenberg 97: 45-55.
Hilgerloh, G. 1989. Autumn migration of trans-Saharan migrating passerines in the straits of Gibraltar. Auk 106: 233-239.
Hilgerloh, G. 1991. Spring migration of passerine trans-Saharan migrants across the straits of Gibraltar. Ardea 79: 57-62.
Houghton, E.W. 1970. Spring Migration at Gibraltar. - RRE Memorandum No 2593.
Kiepenheuer, J. & Linsenmair, K.E. 1965. Vogelzug an der nordafrikanischen Küste von Tunesien bis Rotes Meer. Vogelwarte 23: 80-94.
Laty, M. 1979. Recherches en cours sur les migrations des oiseaux - départs de Camarque au printemps et arrivées de migrateurs sur la c'te méditerranéenne française. Bird Strike Comm. Europe 14/WP7: 1-14.
Liechti, F. & Bruderer, B. 1995. Direction, speed and composition of nocturnal bird migration in the south of Israel. Isr. J. Zool.: 501-515.
Liechti, F., Bruderer, B. & Paproth, H. 1995. Quantification of nocturnal bird migration by moonwatching: comparison with radar and infrared observations. J. Field Ornithol. 66: 457-468.
Liechti, F., Peter, D., Lardelli, R. & Bruderer, B. 1996a. Herbstlicher Vogelzug im Alpenraum nach Mondbeobachtungen - Topographie und Wind beeinflussen den Zugverlauf. Orn. Beob. 93: 131-152.
Liechti, F., Peter, D., Lardelli, R. & Liechti, F. 1996b. Die Alpen, ein Hindernis im nächtlichen Breitfrontzug - eine großrämige Übersicht nach Mondbeobachtungen. J. Ornithol. 137: 337-356.
Liechti, F., Steuri, T., Lopez-Jurado, C., Ribas, P.L.D., Reis, M.A. & Bruderer, B. 1997. Nocturnal spring migration on Mallorca - Schedules of departure and passage. Ardeola 44: 207-213.
Lövei, G.L. 1989. Passerine migration between the Palaearctic and Africa. Current Ornithology 6: 143-174.
Moreau, R.E. 1972. The Palaearctic-African Bird Migration Systems. London and New York: Academic Press.
Nisbet, I.C.T., Evans, P.R. & Feeny, P.P. 1961. Migration from Morocco into Southwest Spain in relation to weather. Ibis 103a: 349-372.
Richardson, W.J. 1978. Timing and amount of bird migration in relation to weather: a review. Oikos 30: 224-272.
Richardson, W.J. 1990. Timing of bird migration in relation to weather: updated review. In Gwinner, E. (ed.) Bird migration: 78-101. Berlin Heidelberg: Springer.
Rivera, C. & Bruderer, B. 1998. Etudes des migrations transméditerranéennes au moyen d'une caméra infrarouge. Directions de vol et topographie régionale. Nos Oiseaux 45, suppl.1: 35-48.
Schüz, E. 1971. Grundriss der Vogelzugkunde. Berlin und Hamburg: Paul Parey.
Spina, F. & Pilastro, A. in press. Strategy of sea and desert crossing in spring passerine migrants as suggested by the analysis of intra- and inter-specific variation of residual fat levels. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1958-1976. Johannesburg: BirdLife South Africa.
Trösch, B., Liechti, F. & Bruderer, B. in press. Do observations of nocturnal autumn migration in southern Spain indicate a concentration towards the Strait of Gibraltar? Ardea.
Wallraff, H.G. & Kiepenheuer, J. 1963. Migracion y orientacion en aves: observaciones en otoño en el sur-oeste de Europa. Ardeola 8: 19-40.
Zink, G. 1973-1985. Der Zug europäischer Singvögel. Ein Atlas der Wiederfunde beringter Vögel., Vol. 1.-4. Lieferung. Möggingen: Vogelzug-Verlag.
Zink, G. & Bairlein, F. 1995. Der Zug europäischer Singvögel. Ein Atlas der Wiederfunde beringter Vögel., Vol. 5. Lieferung. Wiesbaden: Aula.
Fig. 1. Map of suggested directions and concentrations of migratory streams in Europe and the Middle East, primarily based on our own radar, infrared and moon-watch data, bridging the gaps between the punctual information by inference from other sources (e.g. Adams 1962, Kiepenheuer & Linsenmair 1965, Casement 1966, Zink 1973, Alfiya 1995).
Fig. 2. Moon-watch data from various sites during the full-moon phase in September 1996. Data from sites in close neighbourhood are pooled in one circular diagram. The length of the bars in the circular diagrams is proportional to the migration traffic rate MTR in each of twelve sectors. The thickness of the bars indicates the duration of the observations integrated in a diagram (thin < 500 minutes, medium = 500 to 1500 minutes; bold > 1500 minutes), thus providing an index of reliability.
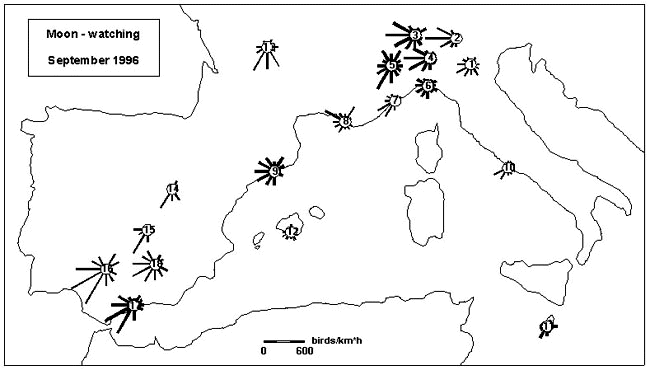
Fig. 3. Moon-watch data from various sites during the full-moon phases in spring 1997. Data from March, April, and May as well as from sites in close neighbourhood are pooled. The length of the bars in the circular diagrams is proportional to the migration traffic rate MTR per sector. The thickness of the bars indicates the duration of the observations integrated in a diagram (as in Fig. 2).
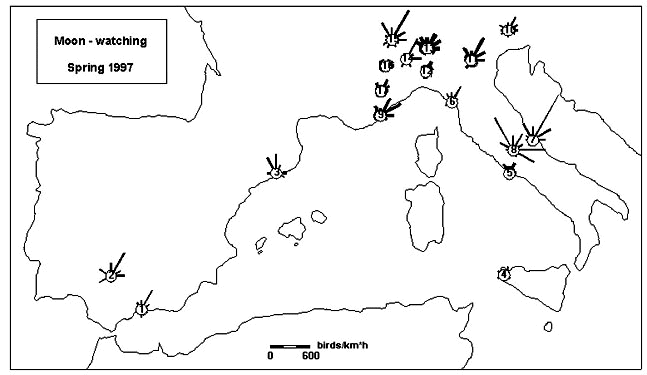
Fig. 4. Seasonal pattern of autumn migration near Malaga.
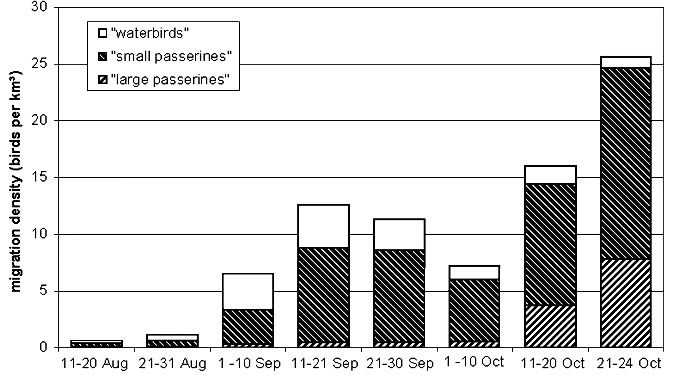
Fig. 5. Diurnal pattern of autumn migration near Malaga. Shading indicates the shifting time of sunrise from August to October.
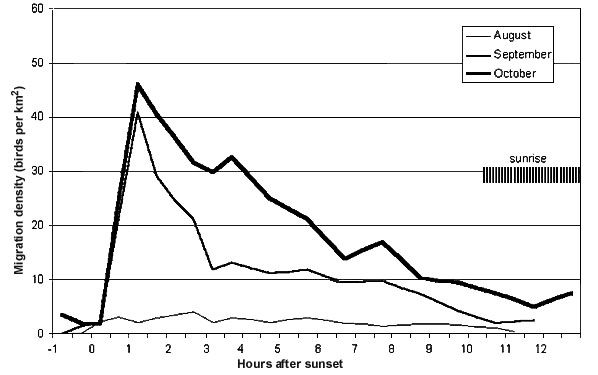
Fig. 6. Diurnal pattern of late autumn migration at the Baleares sites and near Malaga. Shading indicates the shifting time of sunrise from late September to late October.
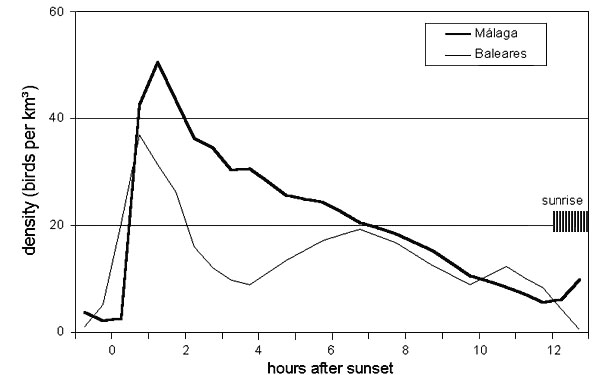
Fig. 7. Seasonal pattern of spring migration at the Malaga (M) and Baleares (B, bold) sites.
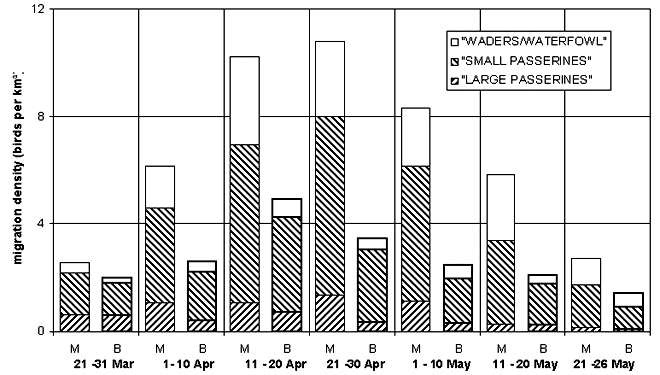
Fig. 8. Diurnal pattern of spring migration near Malaga and at the Baleares site. Shading indicates the shifting time of sunrise from early April to late May.