S33.3: Strategy of sea and desert crossing in spring passerine migrants as suggested by the analysis of intra- and inter-specific variation of residual fat levels
Fernando Spina1 & Andrea Pilastro2
1Istituto Nazionale per la Fauna Selvatica, Via Ca’ Fornacetta 9, I-40064 Ozzano Emilia (BO), Italy, fax 39 51 79 66 28, e-mail infsmigr@iperbole.bologna.it; 2Dipartimento di Biologia, Università di Padova, Via Trieste 75, I-35121 Padova, Italy, e-mail pilastro@civ.unipd.it
Spina, F. & Pilastro, A. 1999. Strategy of sea and desert crossing in spring passerine migrants as suggested by the analysis of intra- and inter-specific variation of residual fat levels. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1958-1976. Johannesburg: BirdLife South Africa.Many islands, scattered in the Mediterranean Sea, are used by passerine migrants for resting during migration. In most cases, migrants do not stay on the islands for refuelling and body condition indexes, therefore, give a good indication of their residual energetic reserves at this stage of barrier crossing. Since 1988, the Progetto Piccole Isole, coordinated by I.N.F.S. as the Italian Ringing Scheme, was aimed at investigating spring migration by a network of 40 insular and coastal stations stretched across the western and central Mediterranean. These stations were operated through standardised mist-netting and ringing methods during the peak migration period of trans-Saharan migrants. Data on body mass, fat and muscle score of over 300,000 birds, belonging to 209 species, were collected during 10 years. These data were used to investigate different aspects of songbird migration on a large geographical scale, in particular: (1) strategy of barrier crossing in terms of routes and phenology; (2) morphological and ecological correlates of residual fat reserves at the inter-specific level; (3) intra-specific variation of body conditions at different stages of sea crossing; (4) intra- and inter-specific patterns of fat and muscle consumption. These results altogether suggest that direct, wide front crossing of the Sahara desert and Mediterranean sea, without intermediate refuelling in North Africa, is a frequent strategy among passerine migrants in spring.
INTRODUCTION
In order to reach southern Europe, spring Palearctic-African migrants are faced with the need to cross two large and inhospitable ecological barriers, represented by the Sahara and the Mediterranean. Passerine migratory strategies over the Sahara have recently started being investigated (Biebach et al. 1986; Bairlein 1987; Biebach et al. 1991; Bairlein 1992; Biebach 1992, 1995; Izhaki & Maitav 1998). The Mediterranean crossing can imply over-flying extensions of open sea of hundreds kilometres. Several authors have pointed out the selective advantage of an early arrival on the breeding grounds, which suggests that migrants could be time-minimising (Alerstam & Lindröm 1990) while moving northwards, and hence more frequently choosing a direct crossing of the barriers.
Prolonged migratory flights across ecological barriers represent extreme situations, which can offer unique opportunities to investigate different aspects of the adaptations evolved to overcome such a delicate and crucial phase of a migrant’s annual cycle. Monitoring bird migration across ecological barriers, and especially wide extensions of sea, is particularly difficult; from this respect, the large number of islands of variable size fairly scattered through the Mediterranean represents an ideal network of field stations.
In general terms, spring migration has been much less studied that the post-nuptial movements, and also many of the proximate and ultimate factors governing the return movements to the breeding grounds have not been thoroughly investigated (Berthold 1993). In order to analyse the different aspects of spring migration across the Mediterranean as an important ecological barrier between Africa and the Palearctic, in 1988 the Italian National Wildlife Institute has launched the ‘Progetto Piccole Isole’ (PPI).
The PPI: aims of the project
The main aims of the project are: (1) to study the strategies adopted, both at the inter- and intra-specific level, by migratory passerines to cross this ecological barrier; (2) to investigate the functional role of islands as stopover sites within these migratory strategies; (3) to monitor return migration in a large number of species, with special emphasis on trans-Saharan migrants, whose populations seem to be significantly limited by ecological factors acting in their African winter quarters (Baillie & Peach 1992).
METHODS
Study period
During the first two pilot years, different phases of the spring migration were investigated (mid-March to mid-April, and mid-April to mid-May in 1989 and 1988 respectively). The later period has been shown to coincide with the peak passage of trans-Saharan migrants (Spina et al. 1993), and therefore a standard field season between 16 April to 15 May has been selected during the following years of the project.
This choice has been a compromise between seasonal and geographical coverage, since a relatively short period has allowed to cover a larger number of sites. Later stages of spring migration have also been tested in some stations (see Montemaggiori et al. 1996).
Ringing activity
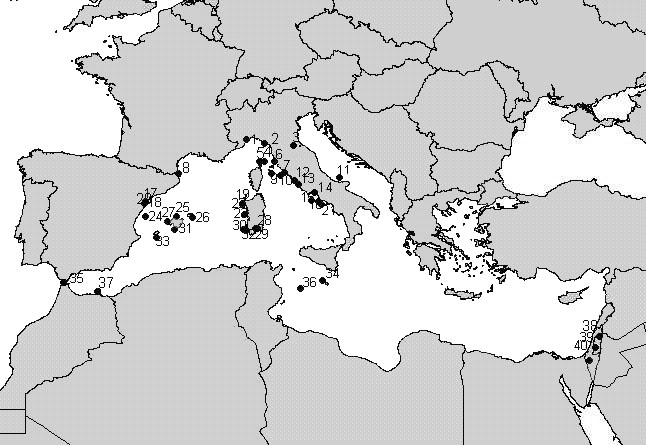
Standardised mist netting was carried on at a series of islands and coastal stations stretched across the Mediterranean. Nets were operated from dawn to dusk, checked every hour and closed only in case of adverse weather conditions. In 11 years of activity, a total of 40 stations (from 7 countries), mainly located in the western and central Mediterranean, have taken part to the project (Fig. 1). Among these, 24 were on islands and 16 on coastal sites. Four stations started the project, and the number of sites which has been involved in the project increased subsequently, with an annual average of 14 stations (range 5-21, Fig. 2). More recently, a few stations located in the eastern Mediterranean have been involved into the project (Fig. 1). Such a co-ordinated ringing effort has only been possible thanks to the contribution of a large number of participants. Over 500 ringers and helpers joined the project, with a positive temporal trend in the annual number of participants (Fig. 3).
Each bird was individually ringed, aged and sexed, and a series of biometrical and physiological data were collected. Wing length was recorded as feather length (Berthold & Friedrich 1979; Jenni & Winkler 1989); birds were weighed to the nearest 0.1g using an electronic balance (Sartorius PT600) or with a Pesola spring balance to the nearest 0.5g. Subcutaneous fat was estimated according to Busse (1974), assigning a score ranging from 0 (no visible fat) to 5 (abdomen and furcula bulging with fat). Most data were computerised at the Italian National Wildlife Institute, where the central data bank derived from the project is hosted.
A total of 397,989 birds have been ringed between 1988-1997, belonging to 209 different species. According also to the number of active stations (see above), a positive temporal trend is recorded both in the annual totals of ringed species (Fig. 4) and individuals (Fig. 5).
RESULTS AND DISCUSSION
To cross or not to cross
One of the first questions is whether birds tend to over-fly the barrier, or rather avoid to do so, by selecting to follow the Mediterranean coasts. From this respect, our network of stations distributed over a wide geographic area offers the opportunity to get an overview of the migratory routes followed by different species.
In order to minimise the potential bias in capture patterns which can be introduced by habitat differences between stations, in an analysis targeted to a number of selected species (Pilastro et al. 1998 in press), the frequency of capture of a given species has been expressed as the frequency relative to the total number of migrants ringed at that station. A sample of 21 stations has been used, where a total of over 74,000 migrants had been ringed. The analysis referred to eight selected species has shown inter-specific differences in the strategy of crossing or not the barrier. In the case of two congeners Hippolais warblers, Melodious Warbler H. polyglotta (Fig. 6) is concentrated along the western Mediterranean coasts, while Icterine Warbler H. icterina (Fig. 7) is very frequent on the stations of the central Mediterranean, indicating how large numbers of birds directly cross the Tyrrhenian sea, along a S-N route. Such routes cannot simply be interpreted as the shortest way from the wintering to the breeding quarters, given the absence of the Melodious Warbler from the central Mediterranean stations, despite the species being a common breeder along the whole Italian peninsula (Meschini & Frugis 1993).
Another very interesting case is represented by Bonelli’s Phylloscopus bonelli and Wood P. sibilatrix Warblers. Also in this case, Bonelli’s Warbler (Fig. 8) seems to prefer to avoid the sea crossing, rather following the western coasts of the Mediterranean, while Wood Warbler (Fig. 9) is represented by large numbers of birds on the central stations of the study area, indicating a direct crossing of the sea. Also in this case, the Italian population of Bonelli’s Warbler seems to reach its breeding grounds from the west and north, rather than directly from the south.
A case of wide front migration across the Mediterranean is offered by the Garden Warbler Sylvia borin, which is a common migrant in all stations (Fig. 10, Grattarola et al. manuscript). The increasing capture frequency with longitude observed on the PPI stations reflects the breeding distribution of the species, which is progressively more abundant in central and eastern Europe (Cramp 1992; Hagemejier & Blair 1997). A significant increase of wing length is observed with longitude (Fig. 11, Grattarola et al. manuscript), matching the west-east cline in size recorded on the breeding grounds of the species (Cramp 1992). Altogether these data suggest that birds from different populations head to their breeding areas along the most direct route, irrespective of the width of sea to be crossed (Bruderer 1994; Bruderer & Liechti 1995). It is therefore clear that some species decide to avoid crossing the barrier, while others do over-fly large stretches of sea; energetic and morphological factors which could help explaining the evolution of such diverse migratory strategies surely deserve further research.
Daily patterns of barrier crossing
Nocturnal migrants confronted with the crossing of ecological barriers like deserts can select an intermittent flight strategy based on stopovers during the day also in inhospitable areas (Biebach 1990; Biebach et al. 1991). When birds have to cross stretches of sea which cannot be overcome during a full night of migration, they may find an island and select to stopover during the day, or otherwise continue to fly. The latitudinal distribution of our ringing sites allows tracking the movements of the fronts of migration across the Mediterranean.
An evidence for the progression of migratory movements during the day across the central Mediterranean has been offered when analysing the daily trapping patterns recorded during three years on 4 islands (Massi et al. 1995). The mean trapping hour shows a clear pattern, with a progressively delayed passage on islands at higher latitudes, as an indication of the fact that birds continue to move north during the day.
A general strategy of a fast crossing of the Mediterranean is confirmed by analyses at the intra-specific level. In the case of the Garden Warbler, a highly significant correlation was found between the number of birds trapped daily on a specific island vs those trapped on three other islands (Pilastro et al. 1995), suggesting that several sites along a S-N route are interested by the a synchronised, wide front of migration.
A progressive northward movement of birds during the day is confirmed by the increasing mean hour of capture with latitude across stations derived from the analysis of a larger sample of over 15,000 Garden Warblers ringed on 18 different stations distributed in the western and central Mediterranean (Fig. 12, Grattarola et al., manuscript). These results indicate that birds continue to fly during the day while on sea crossing; yet, also when landing on an island, their stopover time is very short, as suggested by the very low retrapping rate (A. Pilastro & F. Spina unpubl. results) and the observations of long-distance, nocturnal migrants leaving the islands during daytime (S. Riello & F. Spina unpubl. results).
Physical conditions during the barrier crossing.
When analysing birds which are crossing the barrier, the islands offer the opportunity to investigate energetic aspects of these prolonged flights. It is in particular possible to investigate, both at the inter- and intra-specific level, the average physical conditions observed on islands at different distances from the coasts of North Africa (i.e. at different stages of the sea crossing).
In an analysis carried on at Ventotene (Pilastro & Spina 1997) from a sample of 33,275 birds belonging to 28 species of long-distance migrants, clear inter-specific differences in the average physical conditions were recorded; these patterns show a clear consistency in different years (Fig. 13). A same island seems therefore to be used as a stopover site by species which are represented by large numbers of staging migrants showing variable conditions. In particular the presence at a same site of birds in low, medium and good conditions suggests that not only stranded migrants make use of these stopovers during their barrier crossing.
In this station, situated almost at the end of both the Sahara and Mediterranean crossing, some species still had considerable amounts of fat left, as to allow them to cover hundreds of kilometres, while others had already depleted their reserves, being left with only a very limited residual potential flight range (Fig. 14). When trying to identify the best predictors of the observed residual fat reserves, the only variable found to be correlated was the northernmost latitude of the preferred habitat for the species in Africa (Fig. 15). The group of species with the most critical average conditions were those linked to forest habitats in winter, while the best conditions were shown by those species adapted to open habitats. These results suggest that ecological factors constraint the preparation to migration, as long-distance migrants seem to be forced in their preferred habitats for the fast fattening that precedes the crossing of the Sahara. This is also a clear indication that pre-migratory fuelling takes place not as close as possible to the border of the barrier to overcome (Weber et al. 1998), as the preferred winter habitats can be also quite far from the southern edge of the Sahara.
After the Sahara crossing, birds continue their flight over the Mediterranean apparently without refuelling in North Africa, as shown in over 80% of cases in our group of 28 species, and physical conditions on arrival in southern Europe largely depend on the latitude of the preferred winter habitats south of the Sahara.
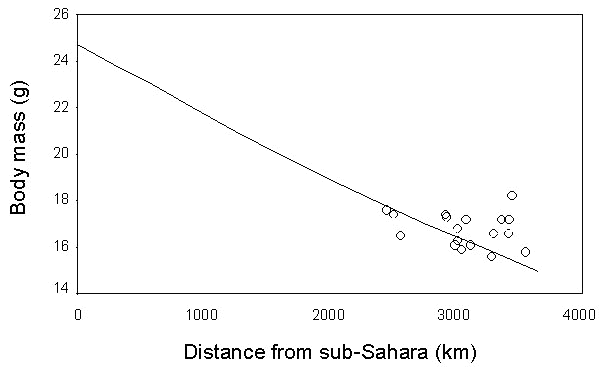
This general strategy recorded in many different species studied in a single station is confirmed also by analyses at the intra-specific level from a large number of sites. In the case of the Garden Warbler (Grattarola et al. manuscript), the geographical variation in the residual fat reserves across the western and central Mediterranean is in agreement with a direct crossing of the desert and sea, as suggested by the observed negative correlation of both fat score (Fig. 16) and body mass (Fig. 17) with increasing latitude. A fast migration without intermediate refuelling is further supported by fat consumption estimates calculated using Pennycuick’s model (1996), which show a clear agreement between the observed and expected values (Fig. 18, Grattarola et al. manuscript).
Birds belonging to different populations of a given species do not seem to differ significantly in their preparation for migration south of the Sahara, as indicated by the absence of a longitudinal pattern in the average physical conditions shown by Garden Warblers migrating across the Mediterranean, despite the observed increase in wing length with longitude (Grattarola et al. manuscript).
Spring migration and the breeding season: differential migration of sex- and age-classes.
The results shown so far indicate a general strategy for a fast spring migration across the Sahara and the Mediterranean. This can be related to the increased speed of migration during barrier crossing (Berthold 1993), but also to a more general feature of spring migration, when birds take much shorter time than in autumn to fly back to their breeding areas from the wintering grounds (Cochran 1987). Such higher average speed during spring migration has been explained by the advantage represented by an early arrival to the breeding grounds, which would increase breeding performances both for males (Dale & Slagsvold 1990, Menzel 1971) and females (Lundberg & Alatalo 1992).
Another peculiarity of spring migration vs. autumn movements is represented by the widespread differential migration of sex- and age-classes (Ketterson & Nolan 1983; Woodrey & Moore 1997; Yong & Moore 1997; Izhaki & Maitav 1998a). Selection acts in favour of such a strategy, as suggested also by the endogenously based differential migratory behaviour described in the case of the Blackcap Sylvia atricapilla (Terrill & Berthold 1989). Although within the Palearctic an earlier arrival of males and adults to the breeding grounds is reported for most species of both medium- and long-distance migrants (Cramp 1985, 1988, 1992; Cramp & Perrins 1993), only scanty information was available on whether this time gap is due to geographic segregation during the winter, to an earlier start of spring migration for males and adults (as in the Yellow Wagtail Motacilla flava, Wood 1992) or to a different speed of migration based also on the differences frequently observed in wing morphology between sex- and age-classes (as in the Willow Warbler Phylloscopus trochilus, Hedenström & Pettersson 1986, or in the Pied Flycatcher Ficedula hypoleuca, Lundberg & Alatalo 1992). In particular, very little attention had been devoted to verify whether sex- and age-classes are already temporally segregated during migration.
In a specific analysis to investigate differential spring migration across the Mediterranean, a group of 16 species of medium- and long-distance migrants ringed on four islands of the central Mediterranean over a period of five years has been taken into account (Spina et al. 1994). A clear temporal gap between both sex- and age-classes has been recorded in most species already during spring migration, and at different stages of barrier crossing. Males have been found passing earlier than females in 14 out of 16 species, apparently irrespective of the location of the species’ main winter quarters in Africa, and despite the inter-annual variability recorded in the general trapping patterns. Age also influences the timing of spring migration, as indicated by a sub-sample of 13 species analysed, where adults have generally been found passing earlier than yearlings in both sexes (18 out of 26 cases).
Competition theory has often been addressed to explain the evolution of migration strategies and sexual segregation (Gauthreaux 1982; Ketterson & Nolan 1983). Differential migration could be explained on the basis of sex-based competition on stopover sites and differential costs of reproduction for the sexes. Ketterson and Nolan (1983) suggest an early arrival on the breeding grounds to be crucially important for males, leading them to winter at more northern latitudes than females, as indicated also by experimental evidence in the Blackcap (Terrill & Berthold 1989). Other studies (Stutchbury 1994) propose that the same pressure acting on the males lead them to select winter habitats which allow an earlier achievement of pre-migratory conditions.
Most long-distance migrants have to overcome at least one ecological barrier during their migration, and in order to do so they store large amounts of energy reserves to sustain long, and often uninterrupted flights. Migrants also have to balance the use of these reserves to satisfy energetic needs both during the fast spring migration and upon arrival on the breeding grounds, where residual fat reserves act as an ‘insurance’ against unfavourable weather conditions and may confer advantages during the breeding cycle (Sandberg 1996, Sandberg & Moore 1996). In the framework of optimal migration theory (Alerstam & Lindström 1990), in a first hypothesis males could be regarded as time-minimisers - being ready to invest a large proportion of their reserves in order to get better quality territories through an early arrival to the breeding grounds - while females might get advantage in trying to reach the breeding grounds still carrying adequate energy reserves, as to be ready to lay eggs shortly after arrival.
The constraints affecting differential migration strategies might act before, during or after the barrier crossing. From this respect, analyses of differences in residual fat loads after crossing an important ecological barrier which offers very limited - if any - opportunities for refuelling, may help investigating the selective pressures modelling differential migration.
In a situation of competition for resources before migration, males would be dominant over females in the fattening areas in Africa, in which case the former would be expected to show higher residual fat levels when reaching the end of the barrier crossing. Since flying efficiency is expected to vary among sexes, a similar situation would be expected if males be regarded as more efficient flyers, given their morphological features (longer and more pointed wings). Alternatively, similar physical conditions would be expected if females could compensate for their lower flying efficiency through higher fat loads at departure.
If the use of energy reserves were modelled by the need of reaching the breeding grounds in good conditions for reproduction, females would be expected to show higher fat residuals. Higher fat loads in females upon arrival on the breeding grounds are reported for a series of both long-, medium- and short-distance migrants (Sandberg 1996).
In order to test among these hypotheses, intra-specific variation of residual fat loads was studied in a sample of 27,136 birds belonging to 11 species of passerines at their arrival on the island of Ventotene, after crossing the Mediterranean (Pilastro et al. unpubl. results). Out of these species, two winter mainly within the Mediterranean, and nine principally or exclusively in trans-Saharan Africa. On the basis of the results of a previous study (Pilastro & Spina 1997), trans-Saharan migrants do not seem to substantially refuel in North Africa, and their conditions on this island can therefore be regarded to as those of birds which have rapidly crossed the desert and sea.
As for sex-based differences in physical conditions, higher residual fat levels have been found in 6 cases for males and 5 cases for females; absolute differences are very small (ca. 1% of the mean body mass), and no significant pattern is observed for sex-classes. These results suggest that if females are less efficient flyers, requiring more energy per unit of distance flown, they do not fully compensate for by storing more reserves before the barrier crossing. Females too seem therefore to try and overcome the Sahara and Mediterranean with a similar strategy as males, i.e. without stops to substantially refuel in North Africa. Differential spring migration of sex-classes seems to be explained as a strategy to reduce intra-specific competition in fattening and stopover sites before embarking on barrier crossing. The sex-based differences in physical conditions observed on the breeding grounds (Sandberg 1996) are therefore likely to originate from later stages of return migration.
CONCLUSIONS
Large numbers of species and birds overcome the barriers represented by the Sahara and Mediterranean during spring migration, often along routes which do not seem to be influenced by the distance to be covered on sea crossing. Return movements are generally fast, and a prevailing time-minimising strategy seems to be adopted by most species of both medium- and long-distance migrants. Differential sex- and age-based migration is a common feature of spring migration strategies; the temporal gap between sex-classes is likely to originate already in the fattening areas south of the Sahara, on the basis of intra-specific competition mechanisms.
The Mediterranean islands represent an important functional component of the Palearctic-African migration system, as they are used for brief stopovers by huge numbers of birds belonging to species with clearly different African origin and destination, and the decision taken by migrants to rest on the islands is not explained by critical physical conditions.
Most species cross both the Sahara and Mediterranean without intermediate refuelling in North Africa. Ecological factors influence the preparation for spring migration, as trans-Saharan migrants are forced into their preferred wintering habitats for storing energy reserves before departure, which addresses conservation problems originating from habitat loss and desertification in Africa.
The Sahara desert and Mediterranean represent an important constraint for Palearctic-African migrants, which seem to make the best of a bad job to overcome these barriers. Integrated conservation policies of the fragile habitats of Mediterranean islands are needed to try and reverse the widespread population decline of many migratory species in Europe (Tucker & Heat 1994). Ongoing research on different aspects of stopover ecology of migrants within the larger ‘Progetto Piccole Isole’ will contribute to the further understanding of selective pressures modelling spring migration across the Mediterranean.
ACKNOWLEDGEMENTS
We wish to thank all ringers who joined the field activities. Without their enthusiasm it would have been impossible to carry out such an intense ringing project. Among them, we are particularly indebted to the coordinators of the ringing stations. For providing permission and support to the project, we wish to thank: Consiglio Nazionale delle Ricerche, Servizio Navi Oceanografiche (Montecristo and Giannutri), Amministrazione Provinciale di Livorno (Capraia), Libero Consorzio di Giannutri e Taverna del Granduca (Giannutri), and Comune di Ventotene. Results from the Progetto Piccole Isole (I.N.F.S.): paper n. 25.
REFERENCES
Alerstam T. & Lindström A. 1990. Optimal bird migration: the relative importance of time, energy and safety. In: Gwinner E. (ed.) Bird Migration: the Physiology and Ecophysiology. Springer Verlag, Berlin: 331-351.
Baillie, S.R. & Peach, W.J. 1992. Population limitation in Palaearctic-African migrant passerines. Ibis 134, suppl. 1: 120-132.
Bairlein, F. 1992. Recent prospects on trans-Saharan migration of songbirds. Ibis, 134, Suppl. 1: 41-46
Berthold, P. 1993. Bird Migration: A General Survey. Oxford; Oxford University Press.
Berthold, P. & Friedrich, W. 1979. Die Federlange. Ein neues nutzliches Flugelmass. Vogelwarte 30: 11-21.
Biebach, H. 1990. Strategies of Trans-Sahara Migrants. In: Gwinner E. (ed.) Bird Migration: the Physiology and Ecophysiology. Berlin; Springer Verlag: 352-367.
Biebach, H. 1992. Flight-range estimates for small trans-Sahara migrants. Ibis 134, Suppl. 1: 47-54.
Biebach, H. 1995. Stopover of migrants flying across the Mediterranean Sea and the Sahara. Israel Journal of Zoology 41 (3): 387-392
Biebach H., Friedrich, W. & Heine, G. 1986. Interaction of bodymass, fat, foraging and stopover period in trans-sahara migrating passerine birds. Oecologia 69: 370-379
Biebach, H., Friedrich, W., Heine, G., Jenni, L., Jenni-Eiermann, S. & Schmidl, D. 1991. The daily pattern of autumn bird migration in the northern Sahara. Ibis 133: 414-422
Bruderer, B. 1994. Radar studies on nocturnal bird migration in the Negev. Ostrich 65: 204-212.
Bruderer, B., & Liechti, F. 1995. Variation in density and height distribution of nocturnal migration in the south of Israel. Israel Journal of Zoology 41: 477-487.
Busse, P. 1974. Biometrical methods. Notatki Orn. 15: 114-126.
Cochran, W.W. 1987. Orientation and other migratory behaviours of a Swainson’s Trush followed for 1500 Km. Animal Behaviour 35: 927-929
Cramp, S. (ed.). 1985. The Birds of the Western Palearctic, Vol. IV. Oxford: Oxford University Press.
Cramp, S. (ed.). 1988. The Birds of the Western Palearctic, Vol. V. Oxford: Oxford University Press.
Cramp, S. (ed.). 1992. The Birds of the Western Palearctic, Vol. VI. Oxford: Oxford University Press.
Cramp, S. & Perrins, C.M. (eds). 1993. The Birds of the Western Palearctic, Vol. VII. Oxford: Oxford University Press.
Dale S. & Slagsvold T. 1990. Random settlement in female Pied Flycatcher, Ficedula hypoleuca: significance of male territory size. Animal Behaviour 39: 231-243.
Gauthreaux, S.A. Jr. 1982. The ecology and evolution of avian migration systems. In: Farner, D.S., King, J.R. & Parkes, K.C. (eds) Avian Biology, Vol. 6, New York; Academic Press Inc.: 93-168.
Hagemeijer, E.J. & Blair, M.J. (eds). 1997. The EBCC Atlas of European Breeding Birds: Their Distribution and Abundance. London; T & AD Poyser.
Hedenström A. & Pettersson J. 1986. Differences in fat deposits and wing pointedness between male and female Willow Warblers caught on spring migration at Ottenby, SE Sweden. Ornis Scandinavica 17: 182-185.
Izhaki, I. & Maitav, A. 1998. Blackcaps Sylvia atricapilla stopping over at the desert edge; physiological state and flight-range estimates. Ibis 140: 223-233.
Izhaki, I. & Maitav, A. 1998a. Blackcaps Sylvia atricapilla stopping over at the desert edge; inter- and intra-sexual differences in spring and autumn migration. Ibis 140: 234-243.
Jenni, L. & Winkler, R. 1989. The feather-length of small passerines: a measurements for wing-length in live birds and museum skins. Bird Study 36: 1-15.
Ketterson, E.D. & Nolan, V. Jr. 1983. The evolution of differential bird migration. In: Johnston, R.F. (ed.) Current Ornithology. Volume 1: 357-402
Lundberg, A. & Alatalo, R.V. 1992. The Pied Flycatcher. London; T. & A.D. Poyser Ltd.
Massi, A., Spina,F. & Montemaggiori, A. 1995. Modalità di attraversamento del Mediterraneo durante la migrazione primaverile. Suppl. Ric. Biol. Selvaggina XXII: 445-451
Menzel, H. 1971. Der Gartenrotschwanz. Wittenberg Lutherstadt.
Meschini, E. & Frugis, S. (eds). 1993. Atlante degli Uccelli nidificanti in Italia. Suppl. Ric. Biol. Selvaggina 20: 1-79.
Montemaggiori, A., Mantovani, R. & Spina, F. (eds). 1996. Progetto Piccole Isole: risultati generali e resoconto del IX anno di attività. Suppl. n. 5 Boll. Attività Inanellamento: 1-91.
Pennycuick, C.J. 1989. Bird flight performance: a practical calculation manual. Oxford: Oxford University Press.
Pennycuick, C.J., Klaasen, M., Kvist, A. & Lindström, A. 1996. Wingbeat frequency and the body drag anomaly: wind-tunnel observations on a thrush nightingale (Luscinia luscinia) and a teal (Anas crecca). Journal of Experimental Biology 199: 2757-2765.
Pilastro, A., Baccetti, N., Massi, A., Montemaggiori, A., Roselli, A. & Spina, F. 1995. Stima della direzione di migrazione e del consumo di grasso per ora di volo nel Beccafiico (Sylvia borin) durante la migrazione primaverile. . Suppl. Ric. Biol. Selvaggina XXII: 453-463.
Pilastro, A. & Spina, F. 1997. Ecological and morphological correlates of residual fat reserves in passerine migrants at their spring arrival in southern Europe. Journal of Avian Biology 28 (4): 309-318.
Pilastro, A., Macchio, S., Massi, A., Montemaggiori, A. & Spina, F. 1998. Spring migratory routes of eight trans-Saharan passerines through the central and western Mediterranean; results from a network of insular and coastal ringing sites. Ibis 140: in press.
Sandberg, R. 1996. Fat reserves of migrating passerines at arrival on the breeding grounds in Swedish Lapland. Ibis 138: 514-524.
Sandberg, R. & Moore, F.R. 1996. Fat stores and arrival on the breeding grounds: reproductive consequences for passerine migrants. Oikos 77 (3): 577-581.
Spina, F., Massi, A., Montemaggiori, A. & Baccetti, N. 1993. Spring migration across central Mediterranean: general results from the ‘Progetto Piccole Isole’. Vogelwarte 37: 1-94.
Spina, F., Massi, A. & Montemaggiori, A. 1994. Back from Africa: who’s running ahead? Aspects of differential migration of sex and age classes in Palearctic-African spring migrants. Ostrich 65: 137-150
Terrill, S.B. & Berthold, P. 1989. Experimental evidence for endogenously programmed differential migration in the blackcap (Sylvia atricapilla). Experientia 45: 207-209.
Tucker, G.M. & Heath, M.F. 1994. Birds in Europe: their conservation status. Cambridge, U.K.: BirdLife International (BirdLife Conservation Series n. 3).
Weber, T.P., Ens, B.J. & Houston, A.I. 1998. Optimal avian migration: a dynamic model of fuel stores and site use. Evolutionary Ecology 12 (4): 377-401.
Wood, B. 1992. Yellow Wagtail Motacilla flava migration from West Africa to Europe: pointers towards a conservation strategy for migrants in passage. Ibis 134 Suppl. 1: 66-76.
Woodrey, M.S. & Moore, F.R. 1997. Age-related differences in the stopover of fall landbird migrants on the coast of Alabama. The Auk 114 (2): 695-707.
Yong, W. & Moore, F.R., 1997. Spring stopover of intercontinental migratory thrushes along the northern coast of the Guld of Mexico. The Auk 114 (2): 263-278.
Fig. 1. Geographical distribution of the PPI project stations in the period 1988-1997. Stations are numbered on decreasing latitude. 1: Bergeggi; 2: Palmaria; 3: Monte Brisighella; 4: Capraia; 5: Capo Corso; 6: Orti Bottagone; 7: Burano; 8: Aiguamolls de l’Empordà; 9: Montecristo; 10: Giannutri; 11: San Domino; 12: Palo Laziale; 13: Castelporziano; 14: Gianola; 15: Ventotene; 16: Vivara; 17: Canal Vell; 18: Encanysada; 19: Punta Cristallo; 20: Punta de la Banya; 21: Capri; 22: Capo Caccia; 23: Maldiventre; 24: Columbretes; 25: Albufera de Alcudia; 26: Illa de l’Aire; 27: Dragonera; 28: Flumini di Quartu; 29: Capo S.Elia; 30: San Pietro; 31: Cabrera; 32: Sant’Antioco; 33: Formentera; 34: Malta; 35: Gibilterra; 36: Lampedusa; 37: Kerbacha; 38: Kfar Rupin; 39: Jerusalem; 40: Sede Boker
Fig. 2. Annual number of PPI project stations in the period 1988-1997
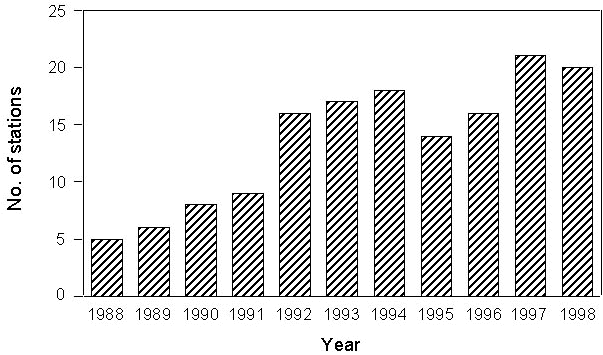
Fig. 3. Annual number of participants to the PPI project in the period 1988-1997
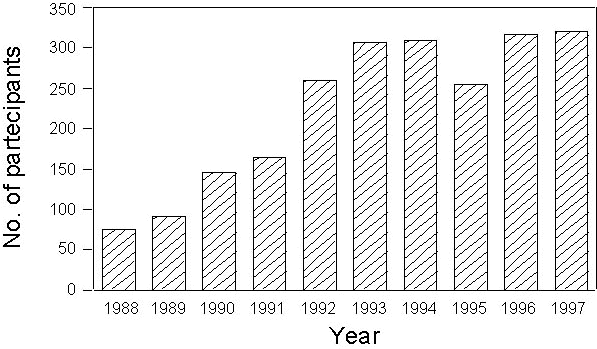
Fig. 4. Annual number of species ringed during the PPI project in the period 1988-1997
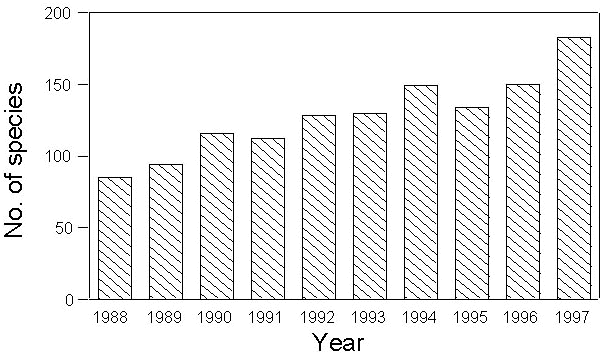
Fig. 5. Annual total number of birds ringed during the PPI project in the period 1988-1997
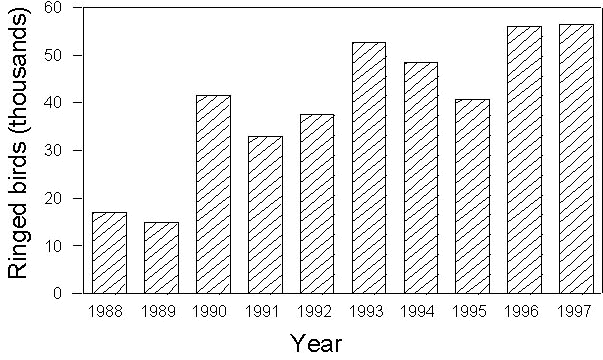
Fig. 6. Capture frequency of Melodious Warbler on 21 PPI stations. Frequencies were standardised in order to vary between 1 for the station with the relative capture frequency (black) and 0 for the station where the migrant was never captured.
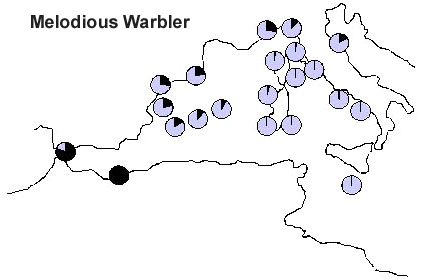
Fig. 7. Capture frequency of Icterine Warbler on 21 PPI stations. Frequencies were standardised in order to vary between 1 for the station with the relative capture frequency (black) and 0 for the station where the migrant was never captured.
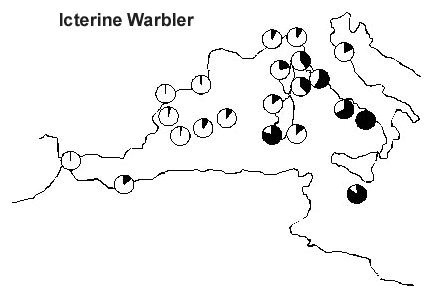
Fig. 8. Capture frequency of Bonelli’s Warbler on 21 PPI stations. Frequencies were standardised in order to vary between 1 for the station with the relative capture frequency (black) and 0 for the station where the migrant was never captured.
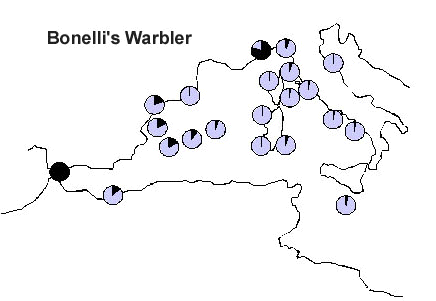
Fig. 9. Capture frequency of Wood Warbler on 21 PPI stations. Frequencies were standardised in order to vary between 1 for the station with the relative capture frequency (black) and 0 for the station where the migrant was never captured.
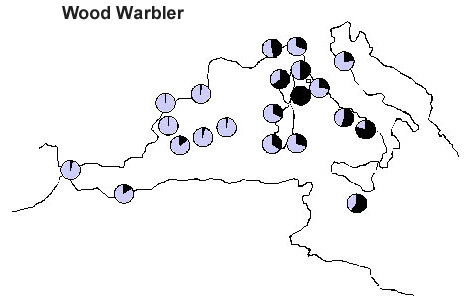
Fig. 10. Capture frequency of Garden Warbler on 18 PPI stations. Capture frequency has been standardised on the number of trans-Saharan migrants captured at each site.
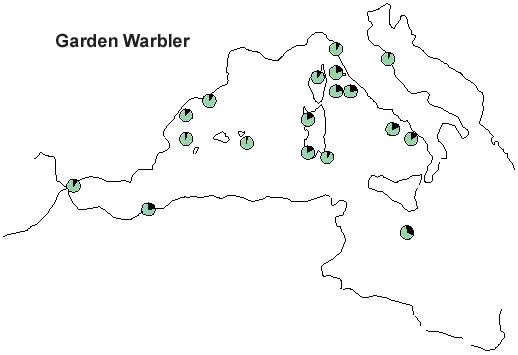
Fig. 11. Mean values of third primary of migrating Garden Warblers in relation to the longitude of the ringing stations. Line is the least square regression fitted to the points (r = 0.89, P<0.001, n = 17).
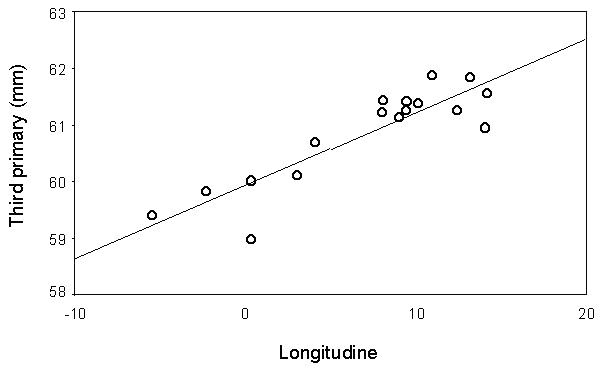
Fig. 12. Mean trapping hour of migrating Garden Warblers in relation to the latitude of the ringing stations. Line is the least square regression fitted to the points (r = 0.58, P = 0.014, n = 17).
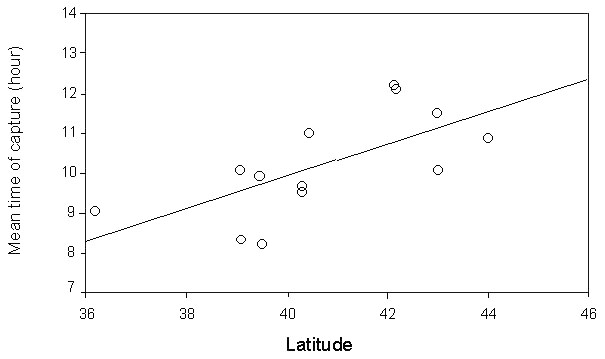
Fig. 13. Mean fat scores of Sedge Warbler Acrocephalus schoenobaenus, Garden Warbler Sylvia borin and Tree Pipit Anthus trivialis recorded for four years on Ventotene island.
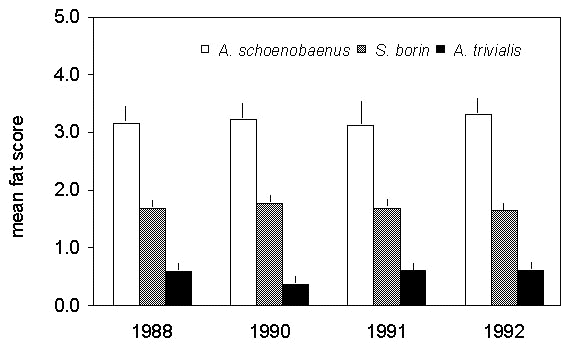
Fig. 14. Residual potential flight ranges of 18 species of long-distance migrants ringed on the island of Ventotene (from Pilastro & Spina 1997).
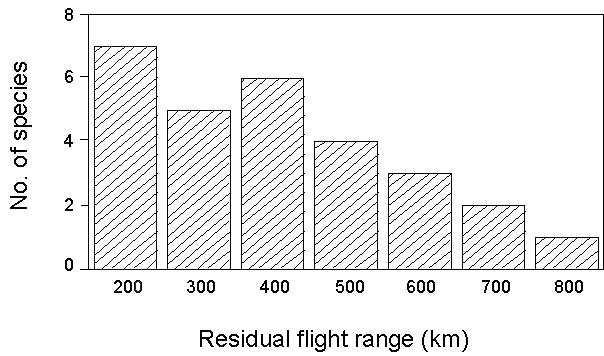
Fig. 15. Mean residual fat load (standardised for the flying efficiency of each species) of 28 long-distance passerine migrants at their arrival on Ventotene island (central Italy). Species are grouped according to their preferred wintering habitats (from Pilastro & Spina 1997).
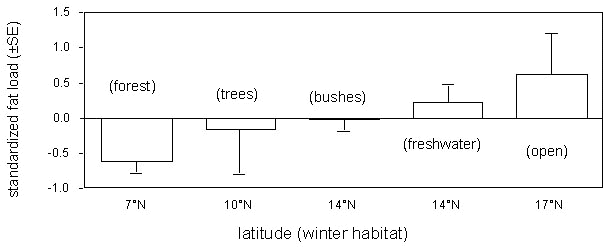
Fig. 16. Mean fat score of migrating Garden Warblers in relation to the latitude of the ringing stations. Line is the least square regression fitted to the points (r= -0.65, P = 0.004, n = 17).
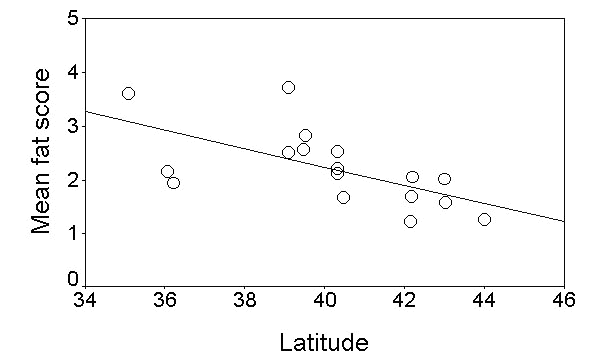
Fig. 17. Mean body mass (corrected for the wing length) of migrating Garden Warblers in relation to the latitude of the ringing stations (only islands). Line is the least square regression fitted to the points (r= -0.67, P = 0.018, n = 12).
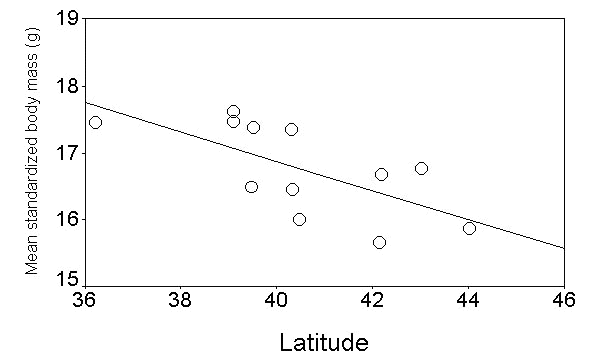
Fig. 18. Observed (points) and predicted (line) body mass values of Garden Warblers at their arrival on the PPI stations. The line represents the expected body mass decrease assuming a take off mass of 25 g, a lean body mass of 14.7 g, and that birds take off at the latitude of 14°N (see Pilastro & Spina 1997). Fat consumption rate was estimated using the version 2 (1997) of the Pennycuick program for flight calculation (Pennycuick 1989; Pennycuick et al., 1996).