S31.5: New developments in bird flight ecology
Adrian L.R. Thomas1& Anders Hedenström2
1Department of Zoology, Oxford University, South Parks Road, Oxford OX1 3PS, UK; *2Department of Animal Ecology, Lund University, Ecology Building, SE-223 62 Lund, Sweden, fax 46 46 222 4716,
anders.hedenstrom@zooekol.lu.se; *address for correspondence Thomas, A.L.R. & Hedenström, A. 1999. New developments in bird flight ecology. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1863-1871. Johannesburg: BirdLife South Africa.Most investigations about bird flight need an understanding of aerodynamics and the power to propel the bird through the air. We have a very useful flight mechanical theory that relates power to airspeed by a U-shaped function. Also for gliding flight, where the energy input comes from potential energy that is converted to kinetic energy rather than muscle work as in flapping flight, there is a closely related functional relationship between rate of sink and forward airspeed. These theories are widely used in various applications of avian ecology, ranging from migration, foraging, escape flight from predator attacks, morphology and the evolution of flight. However, the flight mechanical theory has not been fully validated and it will most likely need further amendments in the light of new experimental data from wind tunnel studies. In spite of this, flight mechanics has inspired a lot of research and it is the basis for the recent developments in optimal bird migration theory and flight behaviour in different ecological contexts. More sophisticated aerodynamic theory has been used for analyses of the evolution of bird tails, where wing theory was instrumental when sorting out the outcome of natural and sexual selection for various tail types. Research in basic aerodynamics is rapidly moving towards very sophisticated methodologies, such as flow visualisation techniques and computational fluid dynamics. Biologists will still gain from interdisciplinary research and we will see a similar development in bird flight research.
INTRODUCTION
The physics of bird flight can be analysed from a fluid mechanics perspective by identifying the different forces acting on the flying bird. Flight theory provides the main tool for predicting expected flight behaviour and optimum morphology of wings, tails and fuselage in birds. Since the 1960ies a quantitative flight mechanical theory has been available, which can be used to calculate power for a given speed on the basis of just a few morphological parameters (e.g. Pennycuick 1968, Tucker 1973). Although this theory is still being tested and refined it has been made available to a wide audience of users, especially avian ecologists, by the publication of easy-to-use computer programs by Pennycuick (1989), and the theory has become known as the Pennycuick theory of bird flight to most ornithologists. The computer programs (for both flapping and gliding flight) encouraged many ornithologists to think about their particular study species in a flight context, and Pennycuick´s book must be one of the most widely cited publications in the ornithological literature. However, in the Pennycuick theory of flapping flight the wing morphology of the bird is represented by its wing span only, which results in wings of very different shape yield the same flight performance if only the span is the same. For more in depth studies of morphological adaptations to flight more a sophisticated wing theory is required (e.g. Jones 1990; Anderson 1991). Such an approach was adopted for detailed studies of the aerodynamics of tail morphology in birds (Thomas 1993), and has recently begun to be applied to gliding (Tucker 1998) and flapping flight (Thomas 1996 a, b), but much more work is needed in this area.
Along another line of research a second layer of theory about optimal migration has been developed during the last decade, by combining basic aerodynamic theory combined with ides from foraging theory (Alerstam & Lindström 1990; Hedenström & Alerstam 1997; Weber et al. 1998). From this development it is evident that aerodynamic principles even dictates behaviour on grounded birds, such as the optimal stopover duration if a bird aims at travelling as fast as possible. This might seem counter-intuitive, but since a bird that accumulates fuel at a stopover increases its weight continuously it also changes its aerodynamic properties. Hence, an extra gram of fat is worth less in terms of potential flight range to a bird that has already accumulated say 50 g of fat than the first gram of fat accumulated by a lean bird. In foraging jargon, accumulated fuel shows a diminishing return function with stopover time, and so the situation is open for trading time at stopover and amount of energy accumulated against each other – optimisation.
However, it may be easy for enthusiastic theoreticians to be carried away and develop theories of limited contact with the real world (see Pennycuick 1998). Therefore, it is important that the basic mechanics of bird flight is confirmed by experiments, such as may be undertaken in a wind-tunnel. But we believe that judicious use of flight theory coupled with comparative morphological analyses or field measurements of flight and migration performance are efficient research strategies. In this paper, we will identify current trends in bird flight research and provide some examples of recent developments.
AERODYNAMICS OF BIRD FLIGHT
A bird in steady flapping flight must generate power to balance both the action of gravity and the drag forces caused by the motion through air. The wings produce lift that counteracts the acceleration due to gravity. The cost of generating the lift force is the induced drag, which decrease with increasing airspeed. Drag is usually decomposed into parasite drag, which is caused by the body (or fuselage) and profile drag, caused by the drag of the wings. Parasite and profile drag are affected by both dynamic pressure and the skin friction. Parasite drag increase with the second power of speed, while profile power is near constant or increase more slowly with speed than parasite drag (Pennycuick 1989). In gliding flight the wings are held in a fixed position to generate lift and the uniform downwards velocity converts kinetic energy into thrust to overcome drag. However, in powered flight the flapping wings must generate lift and thrust. This is achieved by changes in local angle of attack by rotating the wings during a complete wing-beat cycle and by passive aeroelastic mechanisms of the flight feathers, as well as changed span and wing planform between downstroke and upstroke (e.g. Spedding 1992). A complete analytical approach to animal flapping flight that takes account of the changing dynamics during a wing beat is however beyond the current state of science. We have to rely on simplifications and hope that the simple models capture enough of reality to be useful.
Any flight model melts down to the sum of the main three power components (P= DV), and can be expressed as the total mechanical power required to fly P in relation to the flight speed V as
![]()
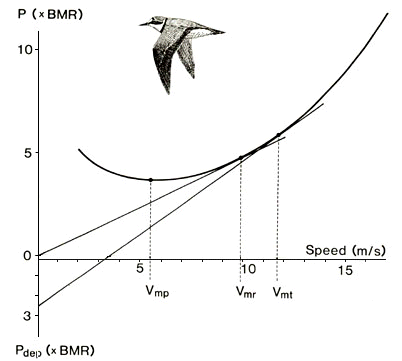
which is the familiar U-shaped power curve (e.g. Pennycuick 1968, 1989). In this equation a represents profile power as constant (although it should probably increase with speed) and b and c represent physical and morphological properties. A typical power curve is shown in Fig. 1. This fundamental relationship between power and airspeed constitutes the backbone of animal flight mechanics.
In principle, it should be straightforward to measure the forces exerted by the pectoralis muscle on the flight bones in the wing, but this is extremely technically challenging as it involves surgical insertion of strain gauges on birds trained to fly in a wind-tunnel. So far, it has only been tried once (Dial et al. 1997), but this study confirmed the general U-shaped relationship between power and speed.
A different approach altogether is to look at the fluid motion in the wake of a flying bird. The impulse imparted to the free-stream airflow is to be found as a deflection downwards of the air passing through the wing disk, and can be found in the form of isolated vortex structures (slow flight) or as an undulating pair of vortex tubes (Spedding 1992). The approach of studying vortex structures has also been used to model the mechanical power of flapping flight (Rayner 1979). That these structures really do exist has been shown qualitatively (Kokshaysky 1979; Spedding et al. 1984; Rayner et al. 1986). By estimating the vorticity in these vortex rings/tubes it is possible to determine mechanical power quantitatively (Spedding et al. 1984; Spedding 1986, 1987). So far this has only been done at a single speed, but with modern digital image velicometry techniques (Fincham & Spedding 1997) there is hope we will be able to accomplish this over a range of speeds and thereby determine a mechanical power curve.
Since the starting point of the analysis of flight was from a physics perspective the theory describes the mechanical power output of the bird, i.e the rate of work imparted on the surrounding air. This is no small achievement, but to generate this power output the bird´s flight muscles must work at a certain frequency to flap the wings. The flight muscles are supplied by energy from metabolic pathways using either carbohydrates, protein or fat as the main substrate. Fat is the most important fuel for migrating birds, but during short flights one of the alternative fuels is probably used. As with all muscular work large quantities of heat are produced as a by-product and this energy cannot be converted into useful mechanical work. In some circumstances it might be diverted to thermoregulation, but generally it is wasted. The conversion efficiency of fuel into mechanical power is probably in the region of 10-20 % (Tucker 1972; Bernstein et al. 1973; Chai & Dudley 1995), so the majority of the power input during flight is not useful aerodynamic power. In addition to aerodynamic power, a real bird also needs to run the normal body functions at a certain metabolic cost and probably also to pay an extra overhead for the increased circulation and respiration associated with flight (Pennycuick 1989). By modifying the mechanical flight theory according to these physiological complications, the mechanical power curve can be converted to
![]()
where r is the physiological overhead during flight, a1 is profile power (a in eqn [1]) and a2 is basal metabolism (Pennycuick 1989). This transformation of the mechanical theory assumes that the conversion efficiency remains constant over the whole speed range, which is perhaps the most parsimonious assumption given the very limited data available. To investigate whether the conversion efficiency really is constant or if it varies in relation to speed should be a research priority in the near future.
To determine the conversion efficiency is important because what matters to the bird when making ecological decisions is the power input or the gross energy cost of flight (which determines how much food the bird needs to find). Then, we will be able to construct a flight theory that is based on mechanics and modified for physiological factors in real birds.
An alternative approach to decide the gross energy cost of flight is to measure the total energy consumption by some physiological method, such as respirometry, doubly labelled water or rate of mass loss. Respirometry has the disadvantages of attaching tubes for collecting exhaled air to the bird, which will affect the aerodynamics and therefore also the results obtained to an unknown degree. Alternatively, the bird can be flown in a sealed wind tunnel where the entire system is a respirometry chamber, but this has the disadvantage of having the bird in a rather confined space with associated problems of aerodynamic interference (e.g. Rae & Pope 1984; Rayner 1994). Estimated flight costs from such methods often show a rather flat relationship between power and speed (Ellington 1991). Although the experimental errors are typically rather large (often larger than the expected deviations of the U-shaped power curve from a horizontal line), some studies show the expected U-shaped relationship (e.g. Rothe et al. 1987). This approach can only be used to construct empirical power curves for particular species and it cannot be generalised in the same way as the flight mechanical theory.
The energy cost of flight is not all that matters. Aerodynamics determine performance of a large set of important flight characteristics, for example the ability to take-off and land, agility, turning radius, acceleration and escape flight during predatory attacks, etc.
OPTIMAL FLIGHT SPEEDS IN ECOLOGICAL CONTEXT
An immediate result from the mechanical theory of flight is the derivation of characteristic flight speeds. Taking the minimum of eqn (1) (dP/dV= 0) gives the flight speed associated with minimum power (Vmp), which is the speed to choose if the objective is to stay airborne for as long time as possible on a given amount of fuel (Fig. 1). If instead minimising P/V (unit energy/ unit distance) we obtain the speed associated with maximum range (Vmr), which is the speed that would bring the bird the longest distance on a given amount of fuel (Fig. 1). The latter speed is the obvious choice to migrating birds if they want to minimise the cost of transport, while Vmp should be selected if the bird intends to maximise its flight time. In which ecological situations can we expect to observe either of these two speeds? Apart from migration, Vmr should be found in birds commuting between food patches or between their nest and feeding sites if they are operating at, or near some metabolic ceiling (Hedenström & Alerstam 1995). A metabolic ceiling is the maximum sustainable rate of energy assimilation (Drent & Daan 1980; Hammond & Diamond 1997). Migrating and breeding birds might well reach such high energy turnover rates (Lindström 1991; Hammond & Diamond 1997).
If birds are minimising the time of migration (sensu Alerstam & Lindström 1990), the associated optimal flight speed (Vmt) is larger than Vmr (Fig. 1). This flight speed depends on the rate at which energy is accumulated at stopovers. With low deposition rates Vmt and Vmr are very close to each other and probably impossible to distinguish in the field given the accuracy of current measurement techniques such as radar and optical range finders.
To measure the flight speed of a bird and compare it with the possible predicted characteristic speeds can be problematic. A recent study of the minimum mechanical power, based on minimum wing-beat frequency in two species flying in a wind-tunnel, suggests that the body drag should be changed to about 0.1 instead of the former default value of 0.4 (Pennycuick et al. 1996). Characteristic flight speeds then become higher than with the old default body drag coefficient. This has led to re-interpretation of observed flight speeds formerly thought of a representing Vmr, but with a lower body drag coefficient (0.1) they come closer to Vmp (Pennycuick 1997). Determining which of the alternative characteristic speeds a particular observed flight speed represents requires a perfect theory of flight and we have not reached that far yet. However, an efficient approach could be to compare the same bird (or at least individuals of the same species) when flying in different situations. Then, at least we know the expected relative expected difference between speeds and situations. One example is the Common Swift Apus apus, which spends the night on the wing when we would expect them to be flying at Vmp in contrast to the expected and higher Vmr during migration. A radar study showed that swifts select flight speed in agreement with predictions (Bruderer & Weitnauer 1972). Another comparison was made in skylarks Alauda arvensis, where the males were expected to select Vmp during sustained song displays and Vmr or Vmt during migration. Also in this case the birds behaved as would be expected from flight mechanics (Hedenström & Alerstam 1996). A third situation could be disorientated birds on migration. When disoriented, which for example could be due to fog, overcast sky or geomagnetic anomalies, a bird should minimise energy consumption by choosing Vmp until orientation clues become available. Emlen & Demong (1978) experimentally released white-throated sparrows Zonotrichia albicollis and used radar to track them when flying off. Under clear skies they flew off at airspeeds very close to their expected Vmr, but when released under overcast conditions, they travelled more slowly at airspeeds close to the expected Vmr as predicted. These comparative studies demonstrate that birds are capable of selecting an appropriate airspeed that may differ substantially between different ecological situations.
It can also be shown that a bird flying at some non- Vmp speed should increase its airspeed into a headwind and reduce it in a tailwind (Pennycuick 1978). That birds do this have been demonstrated repeatedly (e.g. Tucker & Schmidt-Koenig 1971; Bloch & Bruderer 1982; Gudmundsson et al. 1992; Wakeling & Hodgson 1992). For birds flying along a constant track direction it can be shown that they should increase their airspeed in side-winds, independently of the head- or tailwind increment/decrement (Liechti et al. 1994). It remains to show that birds react to this predicted side-wind effect.
During predator attacks it might be wise to disregard energy economics for a moment and just fly as fast or turn as quickly as possible in order to maximise the chances of escaping (Hedenström & Alerstam 1995). Also, the pursuing predator should adjust its airspeed to maximise the chances to capture its quarry, which might not be the maximum possible speed. A recent aerodynamic evaluation showed that large falcons stooping from high altitudes have a capacity of reaching terminal speeds of >100 m/s (Tucker 1998). However, real falcons are generally controlling their speed during stoops at much lower speeds of about 40-50 m/s (Alerstam 1987; Tucker et al. 1998; Peter & Kestenholz 1998).
BEYOND THE U-SHAPED POWER CURVE
The adaptation of birds for flight provides a particularly useful opportunity for the study of evolutionary processes. We can use aerodynamic theory to predict the optimum morphology under selection for particular aspects of flight performance, then test these predictions by using comparative methods. This approach has been successful in studies of bird’s tails (Hedenström 1995), where two new aerodynamic mechanisms have been discovered: an aeroelastic vortex-flap (Norberg 1994) and a novel unsteady control mechanism (Thomas & Balmford 1995). The effects of sexual (Balmford et al. 1993) and natural selection (Thomas & Balmford 1995) on the evolution of the tail have been clarified by using wing theory.
An exciting aim for future research is to extend this approach to bird's wings. There have been valuable attempts to relate wing morphology and flight performance by using fixed wing aerodynamics (Norberg 1990; Spedding 1992), but the basic aerodynamic data (e.g. drag polars, lift coefficients) are lacking. There is, as yet, no theoretical model that can directly relate wing morphology and wing beat kinematics to power output and flight performance (Spedding 1992).
However, recent research has begun to push beyond the simple theoretical analyses underlying the classical U-shaped power curves. Pennycuick (1989), and more recently Tucker (1987, 1998), have considered the effects of the variable geometry of bird’s wings on gliding performance. Because birds can vary their wingspan, wing area, and tail area their gliding performance is not well described by the classical glide polar (an n-shaped parabolic curve relating sink rate and flight speed: the inverse of the U-shaped power curve). Instead they propose the use of a superpolar to describe the performance of gliding birds. The superpolar represents the family of glide polars that characterise a bird’s performance when it adopts a particular geometry in flight. Extending the same logic to powered flight, Thomas (1996 a, b) has shown that rather than a single U-shaped power curve, a bird has a characteristic U-shaped power curve for each wing and tail geometry it can select. Rather than a simple U-shaped curve relating speed and power, when the influence of variable geometry is included the shape of the power curve becomes dependent on the morphology of the bird. For birds with a large forked tail the power curve can be W shaped with two separate local minima: one at low speeds with the tail spread, one at a higher speed with the tail furled.
This approach provides testable predictions of the optimum flight morphology birds should adopt at different flight speeds: If birds experience a U-shaped power curve then to minimise power in flight the wings and tail should be spread widely at low speeds with the tail at a high angle of attack. As speed increases the angle of attack of the tail should be gradually reduced and then completely furled. At high speeds reducing the wing area and wing span reduces power required for flight. These predictions could easily be tested in the field by observing the morphology adopted in steady flight at different speeds.
Computational fluid dynamic models of animal flight are also beginning to appear. Vest & Katz (1996) used an unsteady three dimensional incompressible potential flow model to study the flapping flight of a pigeon flying in a wind-tunnel, and Smith et al. (1996) have advocated the use of unsteady aerodynamic panel methods to analyse flapping flight. However as yet these methods fail to capture the complexity of the motions of birds jointed wings, and the multi-slotted wingtips of most birds would tax computational fluid dynamics (CFD) models even in steady flow. Exciting though CFD is as a way of modelling any particular flying animal, it remains an intensive approach to the analysis of particular problems, and cannot provide the kind of general understanding that analytical models can achieve.
Some of the most recent research has, begun to question whether birds should experience a U-shaped power curve, or even a power curve of any kind. Thomas & Hedenström (1998) point out that as well as a U-shaped relationship between the power required for flight and speed, there should also be some variation in the power available from the muscles with speed. If the power available from the muscles increases with speed, or has some discrete maximum value at some particular speed (as is likely to be the case) then the optimum flight speeds cannot be determined from aerodynamic considerations alone. Some evidence is beginning to suggest that there may indeed be substantial variations with speed in the power available for flight (Rayner & Ward, this volume).
The step beyond the U-shaped power curve has been taken. New theoretical analyses are moving towards the ability to relate flight performance to wing morphology, kinematics, and flight behaviour, and we are at last beginning to see the first rigorous wind tunnel and field testing of the basic elements of flight theory. These are exciting times for research into the aerodynamics and ecology of avian flight. Perhaps soon we will have the theoretical tools that will allow us to directly relate wing morphology and wing beat kinematics to power output and flight performance. The excitement this will trigger stems from the chance, at last, to study the evolutionary processes that have led to that classical example of the perfection that can be achieved through natural selection: the birds wing.
ACKNOWLEDGEMENTS
This work was supported by a Royal Society University Research Fellowship (A. L. R. T.) and the Swedish Natural Science Research Council (A.H.).
REFERENCES
Alerstam, T. 1987. Radar observations of the stoops of the peregrine falcon Falco peregrinus and the goshawk Accipiter gentilis. Ibis 129: 267-273.
Alerstam, T. & Lindström, Å. 1990. Optimal bird migration: the relative importance of time, energy, and safety. In: Gwinner, E. (ed.), Bird migration: physiology and ecophysiology; Berlin, Springer: 331-351.
Anderson, J.D., Jr. 1991. Fundamentals of aerodynamics. 2nd edn. New York; McGraw-Hill: 772 pp.
Balmford, A., Thomas, A.L.R. & Jones, I.L. 1993. Aerodynamics and the evolution of long tails in birds. Nature, Lond. 361: 628-630.
Bernstein, M.H., Thomas, S.P. & Schmidt-Nielsen, K. 1973. Power input during flight of the fish crow, Corvus ossifragus. J. exp. Biol. 58: 401-410.
Bloch, R. & Bruderer, B. 1982. The air speed of migrating birds and its relationship to the wind. Behav. Ecol. Sociobiol. 11: 19-24.
Bruderer, B. & Weitnauer, E. 1972. Radarbeobachtungen über Zug und Nachtflüge des Mauerseglers (Apus apus). Rev. Suisse Zool. 79: 1190-1200.
Chai, P. & Dudley, R. 1995. Limits to vertebrate locomotor energetics suggested by hummingbirds hovering in heliox. Nature, Lond. 377: 722-725.
Dial, K.P., Biewener, A.A., Tobalske, B.W. & Warrick, D.R. 1997. Mechanical power output of bird flight. Nature, Lond. 390: 67-70
Drent, , R.H. & Daan, S. 1980. The prudent parent: energetic adjustments in avian breeding. Ardea 68: 225-252.
Ellington, C.P. 1991. Limitations on animal flight performance. J. exp. Biol. 160: 71-91.
Emlen, S.T. & Demong, N.J. 1978. Orientation strategies used by free-flying bird migrants: a radar tracking study. In: Schmidt-Koenig, K. & Keeton, W. T. (eds), Animal migration, navigation, and homing. Berlin; Springer-Verlag: pp. 283-293.
Fincham, A.M. & Spedding, G.R. 1997. Low cost, high resolution DPIV for measurement of turbulent fluid flow. Experiments in Fluids 23: 449-462.
Gudmundsson, G.A., Alerstam, T. & Larsson, B. 1992. Radar observations of northbound migration of the Arctic tern, Sterna paradisaea. At the Antarctic Peninsula. Antarctic Science, Wash. 4: 163-170.
Hammond, K.A. & Diamond, J. 1997. Maximal sustained energy budgets in humans and animals. Nature, Lond. 386: 475-462.
Hedenström, A. 1995. Swallows unhandicapped by long tails? Trends in Ecology & Evolution 10: 140-141.
Hedenström, A. & Alerstam, T. 1995. Optimal flight speed of birds. Phil. Trans. Roy. Soc. Lond. B 348: 471-487.
Hedenström, A. & Alerstam, T. 1996. Skylark optimal flight speeds for flying nowhere and somewhere. Behav. Ecol. 7: 121-126.
Hedenström, A. & Alerstam, T. 1997. Optimum fuel loads in migratory birds: distinguishing between time and energy minimisation. J. theor. Biol. 189: 227-234.
Jones, R.T. 1990. Wing theory. Princeton; Princeton University Press: 216 pp.
Kokshaysky, N.V. 1979. Tracing the wake of a flying bird. Nature, Lond. 279: 146-148.
Liechti, F., Hedenström, A. & Alerstam, T. 1994. Effects of sidewind on optimal flight speed of birds. J. theor. Biol. 170: 219-225.
Lindström, Å. 1991. Maximum fat deposition rates in migrating birds. Ornis Scand. 22: 12-19.
Norberg, R.Å. 1994. Swallow tail streamer is a mechanical device for self-deflection of tail leading edge, enhancing aerodynamic efficiency and flight manoeuvrability. Proc. Roy. Soc. Lond. B 257: 227-233.
Norberg, U.M. 1990. Vertebrate Flight. Berlin; Springer-Verlag: 291 pp.
Pennycuick, C.J. 1968. Power requirements for horizontal flight in the pigeon Columba livia. J. exp. Biol. 49: 527-555.
Pennycuick, C.J. 1978. Fifteen testable predictions about bird flight. Oikos 30: 165-176.
Pennycuick, C.J. 1989. Bird flight performance: a practical calculation manual. Oxford; Oxford University Press: 153 pp.
Pennycuick, C.J. 1997. Actual and ‘optimum’ flight speeds: field data reassessed. J. exp. Biology. 200: 2355-2361.
Pennycuick, C.J. 1998. Towards an optimal strategy for bird flight research. J. Avian Biol. 29: 449-457
Pennycuick, C.J., Klaassen, M., Kvist, A. & Lindström, Å. 1996. Wingbeat frequency and the body drag anomaly: wind-tunnel observations on a Thrush Nightingale (Luscinia luscinia) and a teal (Anas crecca). J. exp. Biol. 199: 2757-2765.
Peter, D. & Kestenholz, M. 1998. Stuzflüge von Wanderfalke Falco peregrinus und Wüstenflake F. pelegrinoides. Orn. Beob. 95: 107-112.
Rae, W.H. & Pope, A. 1984. Low-speed Wind Tunnel Testing, 2nd edn. New York; John Wiley & Sons: 534 pp.
Rayner, J.M.V. 1979. A new approach to animal flight mechanics. J. exp. Biol. 80: 17-54.
Rayner, J.M.V. 1994. Aerodynamic corrections for the flight of birds and bats in wind tunnels. J. zool., Lond. 234: 537-563.
Rayner, J.M.V., Jones, G. & Thomas, A.L.R. 1986. Vortex flow visualisations reveal change of upstroke function with flight speed in microchiropteran bats. Nature, Lond. 321: 162-164.
Rothe, H.-J., Biesel, W. & Nachtigall, W. 1987. Pigeon flight in a wind tunnel. II. Gas exchange and power requirements. J. Comp. Physiol. B 157: 99-109.
Smith, M.J.C., Wilkin, P.J. & Williams, M.H. 1996. The advantages of an unsteady panel method in modelling the aerodynamic forces on rigid flapping wings. J. exp. Biol. 199: 1073-1083.
Spedding, G.R. 1986. The wake of a jackdaw (Corvus monedula) in slow flight. J. exp. Biol. 125: 287-307.
Spedding, G.R. 1987. The wake of a kestrel (Falco tinnunculus) in flapping flight. J. exp. Biol. 127: 59-78.
Spedding, G.R. 1992. The aerodynamics of flight. In: Alexander, R. McN. (ed.) Mechanics of Animal Locomotion. Adv. Comp. Environ. Physiol. 5: 51-111.
Spedding, G.R., Rayner, J.M.V. & Pennycuick, C.J. 1984. Momentum and energy in the wake of a pigeon (Columba livia) in slow flight. J. exp. Biol. 111: 81-102.
Thomas, A.L.R. 1993. On the aerodynamics of birds´tails. Phil. Trans. Roy. Soc. Lond. B 340: 361-380.
Thomas, A.L.R. 1996a. The flight of birds that have wings and a tail: variable geometry expands the envelope of flight performance. J. theor. Biol. 183: 237-245.
Thomas, A.L.R. 1996b. Why do birds have tails? The tail as a drag reducing flap and trim control. J. theor. Biol. 183: 247-253.
Thomas, A.L.R. & Balmford, A. 1995. How natural selection shapes birds’ tails. Am. Nat. 146: 848-868.
Thomas, A.L.R. & Hedenström, A. 1998. The optimum flight speeds of flying animals. J. Avian Biol. 29: 469-477.
Tucker, V.A. 1972. Metabolism during flight in the laughing gull, Larus atricilla. Am. J. Physiol. 222: 237-245.
Tucker, V.A. 1973. Bird metabolism during flight: evaluation of a theory. J. exp. Biol. 58: 689-709.
Tucker, V.A. 1987. Gliding birds: the effect of variable wing span. J. exp. Biol. 133: 33-58.
Tucker, V.A. 1998. Gliding flight: speed and acceleration of ideal falcons during diving and pull out. J. exp. Biol. 201: 403-414.
Tucker, V.A., Cade, T.J. & Tucker, A.E. 1998. Diving speeds and angles of a gyrfalcon (Falco rusticolus). J. exp. Biol. 201: 2061-2070.
Tucker, V.A. & Schmidt-Koening, K. 1971. Flight speeds of birds in relation to energetics and wind directions. Auk 88: 97-107.
Wakeling, J.M. & Hodgson, J. 1992. Optimisation of the flight speed of the little, common and sandwich tern. J. exp. Biol. 169: 261-266.
Weber, T.P., Ens, B.J. & Houston, A.I. 1998. Optimal avian migration: A dynamic model of fuel stores and site use. Evol. Ecol. 12: 377-401.
Vest, M.S. & Katz, J. 1996. Unsteady aerodynamic model of flapping wings. AIAA Journal 34: 1435-1440.
Fig. 1. The relationship between power and speed in flapping flight as calculated from flight mechanical theory showing the typical U-shaped function. Power and rate of energy deposition of migrating birds when at stopovers (Pdep; negative ordinate) have been related to basal metabolic rate for the bird. Also indicated three characteristic flight speeds of ecological significance: minimum power speed (Vmp), maximum range speed (Vmr) and the optimal flight speed associated with maximum overall migration speed (Vmt).