S31.3: The consequences of flight for the evolution of tail ornaments in birds.
Matthew R. Evans
Department Biological and Molecular Sciences, University Stirling, Stirling, FK9 4LA, UK, e-mail MRE2@stir.ac.uk
Evans, M.R. 1999. The consequences of flight for the evolution of tail ornaments in birds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1823-1843. Johannesburg: BirdLife South Africa.Males of many bird species have elongated tails which have been shown to play a role in mate choice. The possession of an elongated tail has aerodynamic and mechanical consequences for a bird. These consequences will vary with tail shape and length. A model of bird flight was produced in which tail shape and length were varied. When only a small number of tail feathers are elongated the dominant effect was the Norberg mechanism which increases lift and reduces drag and power for forked tails. Whatever the tail shape, when all tail feathers are elongated the delta wing effect which increased lift (if tails are stiff), drag and power become more important than the Norberg effect. Turn radii in gliding and flapping flight are influenced mainly by the Norberg mechanism reducing turn radius in forked tailed birds. The power required at minimum power speed illustrates that for small birds there are large regions of tail space that are unavailable as the cost of flight exceeds the available power. This will have large consequences for ornament evolution in some species.
INTRODUCTION
Males of many bird species have elongated tails. The role of male tail length has been investigated in several species and a relationship between male tail length and female choice has been shown in a number of studies (for review see Andersson 1994). However, even in species which have tail ornaments, the tail of a bird still plays an important role in flight. The most important aerodynamic roles played by the tails of birds seem to be during manoeuvres and in slow flight.
Using slender lifting surface theory Thomas (1993a) was able to show that the tail of a bird can generate substantial lift when held at small angles of attack to the incident airflow. Lift generated by the tail is likely to be important when the bird is in slow flight and during manoeuvres. It is during these types of flight that birds tails are typically spread (Rayner 1987). Thomas (1993a) calculated that the lift generated by a bird's tail is proportional to the square of the maximum continuous span of the tail.
Most of the lift of a bird’s tail is generated by the leading edges, formed by the outermost tail feathers. A bird’s tail approximates to a delta wing and it is known that if the leading edges of a delta wing are drooped, the lift acting on the leading edges is increased and the drag reduced (Lee & Ho 1990; Thomas 1993a). Norberg (1994) suggested that the elongated tail streamers seen in some forked tailed birds could be automatic control devices that deflect the leading edges of the tail to form a vortex flap whenever the tail is at an angle to the incident airflow. Theoretically this mechanism can reduce drag by about 20% and increase the lift to drag ratio by 30%. However, it is at yet unknown whether this mechanism actually occurs when birds use their tails.
It is clear that over the last five years aerodynamic theory as applied to birds tails has developed considerably. However, there has been no attempt to apply these new aerodynamic ideas to explain the diversity of birds tails seen in nature. Evans & Thomas (1992) considered a bird’s tail as a flat plate which produced only drag. This was modelled by modifying the standard Pennycuick (1975, 1989) equations. It is now clear that the tail can also act as a lift producing surface which was how Thomas (1993a, 1996) considered the tail. Thomas (1993a, 1996) used a combination of Pennycuick’s (1975, 1989) momentum jet model and his own delta wing theory. Thomas (1993a) considered that all tail elongation posterior to the maximum continuous span could only produce drag. This meant that forked tails produced the same lift as graduated tails provided that the maximum span and length were the same. The advent of Norberg’s (1994) ideas means that forked tails will provide quantitatively different aerodynamic parameters from graduated tails even if they are the same size as the streamers will act to increase lift and reduce drag. One objective of this paper is to present a model that encompasses all these aspects of theory.
Clearly the fact that the tail of a bird has to perform an aerodynamic role(s) will conflict and interact with its signalling function in some species. It also seems intuitive that the requirement for aerodynamic function could determine whether the tail could be used for advertisement in some species and could also influence the type and extent of tail elongation. In support of this it has been shown (Balmford et al. 1993) that graduated tails (in which all tail feathers are elongated) are more common in families of birds which do not rely on flight frequently. Conversely, families in which flight is an important part of their ecology tend to have streamer tails (in which only the outer tail feathers are elongated). The second objective of this paper is to apply theory in an attempt to explain some of the diversity of tail shapes and lengths seen in different species of birds.
There are several aerodynamic and mechanical parameters that could be influenced by the development of an elongated tail (Evans & Thomas 1992).
Body mass
The mass of the tail contributes to the body mass of the bird and this has a direct effect on the cost of flight. All models of flight agree that at low speeds and in hovering the power required to support weight dominates power output, at least in vertebrates (Pennycuick 1975; Rayner 1979a,b, 1987; however see also Dudley & Ellington 1990). Adding mass to a bird would increase the cost of flight, particularly at low speeds, and would affect the optimum airspeeds for minimum power and for maximum range. The maximum range of the bird and the cost of transport (energy consumption per unit distance travelled) would also be affected (Pennycuick 1975; Rayner 1987).
Changes in body mass have also been shown to affect agility and manoeuvrability (Aldridge 1986; Hughes 1990). Two effects are important here; firstly there is a direct reduction of the power to weight ratio, important in grunt manoeuvres (initiating rapid climbs, accelerating, stopping, etc.). Secondly the force which must be provided by the wings to turn the bird is dependent on the mass of the bird, and both agility and manoeuvrability are in the end limited by the maximum lift coefficient of the wings and by the strength of the wing structures.
Static balance
Adding mass to the tail affects the balance of the bird by adding a turning moment. This may be more costly than adding mass at the centre of gravity of the bird. The magnitude of the effect will be directly proportional to the mass added times the distance between the centre of gravity of the bird and the centre of gravity of the added mass (see Alexander 1983).
Moment of inertia of the animal
The moment of inertia of a body is a measure of its resistance to rotational acceleration and is proportional to the product of the mass of the body and the radius squared. Adding a long tail will increase the moment of inertia of the entire animal reducing agility. If the mass of the tail amounts to a significant proportion of the mass of the animal this will be a significant effect.
Moment of inertia of the tail: stability and control
The tail of the bird is used as a trim and control surface and is rapidly and continuously moved during flight both to maintain stability and to induce turns about all three axes of the bird. The tail is the main control surface relevant to pitch in a bird, and a bird without a tail is barely able to fly due largely to pitch instability. The tail is also of great importance in the control of yaw and roll during turns, however there is insufficient data to enable these factors to be adequately explored (see for example Hertzog 1968; King & King 1979; Bilo et al. 1985).
Adding mass to the tail will increase its moment of inertia directly affecting the force needed to move the tail relative to the bird. The moment of inertia of the tail is proportional to its mass times the square of the distance from the centre of gravity of the tail to its pivot point. Because the length term is squared and because the tail is constantly accelerated to effect control this will be a large effect in any animal with an elongated tail. This method gives a simple estimate of the maximum moment of inertia of the tail and assumes the tail to be a stiff rod. The birds can reduce the moment of inertia by making the tail flexible. Evans & Thomas (1992) showed that this was one of the dominant effects of tail length manipulation in Scarlet-tufted Malachite Sunbirds Nectarinia johnstoni and could explain the reduction in hawking efficiency seen in this species after tail elongation.
Lift and drag: cost of flight
Altering the length of the tail would change its area and possibly its maximum continuous span. The contribution of lift generated by the tail is an important factor in manoeuvrability; manoeuvrable animals have larger wing surface areas and larger tail areas than average. The disadvantage of a large tail is that drag is proportional to area and the drag of the tail is a significant contribution to the parasite drag of the animal (the drag of the body). The normal tail of a bird may contribute as much as 10% of the total drag at high speeds (this is why birds fold their tails during steady flight) (Rayner 1987). At the normal cruising speeds the power to overcome drag is the dominant cost of flight.
Turn radius
The minimum turn radius that can be achieved by a bird depends on the maximum lift coefficient it can generate. During a turn, lift balances weight and also provides the turning force. The lift produced by the bird’s tail is likely to be particularly important during turning flight as the tail is spread during turns.
Aerodynamic stability
A long tail will act like the feathers on a dart and tend to stabilise flight passively. A stable aircraft tends to fly in a straight line. To initiate any turn a force must be applied so stability reduces agility. Birds, like fighter aircraft, are able to exploit the agility that comes with an unstable design by actively maintaining stability through continuous control movements. The aerodynamic stability of animals remains a neglected topic, in the absence of any solid information about this effect any further consideration would be speculation
METHODS
Classification of tail shapes
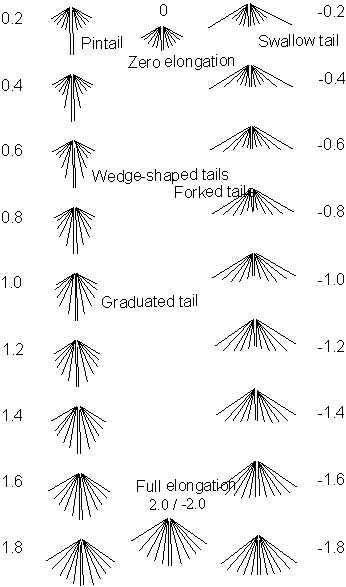
One objective of this paper is to apply current aerodynamic theory to a range of tail shapes. In order to do this tail shapes were classified on a continuous scale from -2 to 2 (Fig. 1). The scale is symmetrical with zero representing a tail in which all the feathers are of equal length and unelongated, 0.2 represents a pintail in which the central pair of rectrices are elongated and the rest are unaltered. As tail shape moves from 0.2 to 1.0 a larger proportion of the feather are elongated, such that 0.4 represents a tail in which the inner pair are fully elongated and the 2nd pair are elongated to half the length of the inner pair, 1.0 represents a tail with all tail feathers except the outer pair are elongated with an even distribution of elongations between zero and full elongation as you move from the outside edge to the centre. Between 1.0 and 2.0 the outer feather elongate from zero elongation at 1.0 to full elongation at 2.0, the inner pair of feathers are always fully elongated, the intermediate feathers have evenly distributed elongations between the outer feather length and the inner feather length. At 2.0 all the feathers are the same length at full elongation. The negative side of the scale represents a similar set of tail shapes but starting with elongation of the outer pair of feathers.
This classification means that a sunbird’s (e.g. Nectarinia johnstoni) tail would score 0.2, a swallow’s (e.g. Hirundo rustica) -0.2, a pheasant (Phasianus colchius) 1.0, a kite (e.g. Milvus milvus) -1.0.
The model considers tail shapes at intervals of 0.2 between 2 and -2. Tail elongations are varied from zero to one metre.
Mechanical parameters
Mechanical parameters are those which impact upon the bird irrespective of whether or not it is in flight. As described above, four mechanical effects of tail elongation are considered
Body mass
Tail mass was calculated as the change in tail area multiplied by the mass per unit area of feather (0.0073 kgm-2). Tail mass was converted to a percentage of body mass to examine the impact of changing tail mass.
Static balance
The turning moment of the tail was calculated as the mass of the tail multiplied by the acceleration due to gravity multiplied by the lever arm.
Moment of inertia of the animal
This was calculated assuming that the bird was a homogeneous cylinder. The moment of inertia of a cylinder relative to its centre of gravity is proportional to its mass, radius squared and height squared.
The moment of inertia of the elongated tail around the bird’s centre of gravity was calculated by adding the mass of the elongated feathers at a distance calculated as the distance between tail’s centre of gravity and the bird’s centre of gravity. The percentage change in total moment of inertia was calculated.
Moment of inertia of the tail: stability and control
The moment of inertia of the tail pitching as a unit around the point at which the tail attaches to the body was calculated as the mass of the tail multiplied by the distance between the tail centre of gravity and the point at which the tail is attached to the body.
Aerodynamic Parameters
Aerodynamic parameters only impact upon the bird during flight. Four aerodynamic effects of tail elongation are considered.
Lift
The total lift is calculated as the sum of the lift produced by the wings and the tail. The lift produced by the wings is proportional to wing area and the square of air speed (Pennycuick 1975, 1989). Tail lift is calculated following Thomas (1993a) as proportional to the angle of attack of the tail, the square of the maximum continuous span and the square of air speed. The Norberg effect is included by the use of a lift enhancement factor which multiplies the lift produced by the tail by 30% when the outer tail feathers 120 mm long and by proportionately more or less if the outer tail feathers are longer or shorter than that. The rationale for this is that the strength of this effect has only been estimated in swallows (A.L.R. Thomas personal communication) in which a lift enhancement of 30% was produced for a model with 120mm long streamers. The Norberg effect is assumed to operate only on the outer tail feathers and only on that part which projects beyond the second outermost feathers.
Drag
The total drag on the bird is calculated as the sum of induced drag, profile drag and parasite drag on the bird’s body and wings (Pennycuick 1975, 1989) and the tail drag. The drag produced by the bird’s tail is the sum of the tail induced drag and the tail profile drag multiplied by a Norberg effect drag reduction factor. Tail induced drag is proportional to tail lift and the angle of attack, profile drag is proportional to tail area and the square of air speed (Thomas 1993a). The Norberg drag reduction factor is used in a similar manner to the lift enhancement factor. Drag is reduced by 50% when the length of the outer tail feather projecting beyond the second outermost is 120mm and by proportionately more or less if the outer tail feathers are longer or shorter respectively. These figures are based on estimates of the strength of the effect in swallows (A.L.R. Thomas personal communication)
Power
Power requirements were calculated using Pennycuick’s standard model of animal flight (Pennycuick 1989). In this model power is proportional to air speed and total drag. Maximum range speed was estimated as the tangent (passing through the origin) to the airspeed v power curve, minimum power speed as the minimum of the airspeed v power curve. The maximum power which could be produced by a bird was estimated by assuming the flight muscles comprised 25% of body mass and generated 100 Wkg -1 (Marden 1994). Power was calculated for level flight (tail furled, 0° tail angle of attack, no Norberg effect) and turning flight (tail spread to 120°, 5° tail angle of attack, Norberg effect operating).
Turn radius
Turn radius was calculated following Thomas (1993b). Turn radii can be calculated for gliding flight (neglecting drag, i.e. assuming no loss of altitude) and flapping flight (including drag). Minimum turn radius is proportional to the square of airspeed and the square of body mass and inversely proportional to the square of total lift. For flapping flight minimum turn radius is also inversely proportional to the square of total drag.
RESULTS
Mechanical parameters
Body mass
Tail mass changes relatively little, tails have to be large with many elongated feathers before tail mass impacts strongly as a percentage of body mass (Fig. 2). Obviously this varies with the size of the bird considered but even a 10 g bird would only have 20% of its body mass represented by a 1m long graduated tail.
Static balance
The turning moment of the tail is very small when only a few feathers are elongated but appears to increase rapidly as all the feathers start elongating (Fig. 3). Large tails can have high turning moments and potentially could influence static balance significantly for small birds.
Moment of inertia of the animal
The moment of inertia of the animal increases rapidly with increases in tail length and shape. Fig. 4 shows the percentage of the bird’s moment of inertia provided by the tail in a 10 g bird. Long tails with many elongated feathers represent a very high proportion of the bird’s moment of inertia. This will significantly reduce agility (agility decreases in direct proportion to the bird’s moment of inertia). This will be reduced if the tail is not stiff.
Moment of inertia of the tail: stability and control
The moment of inertia of the tail increases rapidly as tails get longer and have a greater number of elongated feathers (Fig. 5). This will have a significant impact on control and agility. The bird’s ability to manoeuvre will be reduced and its perceived risk of crashing will probably increase.
Aerodynamic Parameters
All the aerodynamic parameters considered vary with airspeed. This means that the model output is in the form of a four-dimensional figure. The figures used here are three-dimensional cross sections through this four-dimensional figure.
Lift
There are three main effects that can be seen in the lift surface.
(1) At low airspeeds there is very little effect of either shape or length, at high airspeeds both have strong effects, this can be seen by comparing the graph for tail shape and length against lift at 5 ms-1 and at 14 ms-1 (Fig. 6)
(2) The Norberg effect can be seen to enhance lift particularly at high airspeeds for swallow tailed birds (shape = -0.2) and generally increases lift for all forked tailed birds (shape = -0.2 to -1.0) (Fig. 6)
(3) The delta wing effect has a large impact on tails in which all the feathers are elongated (-2 to -1 and 1 to 2). It is of course only in this region that the maximum continuous span starts to increase as it is largely set by the angle of spread and the length of the shortest feathers. The delta wing effect is very large compared to the Norberg effect. For example, once all the tail feathers elongate and increase of only 100 mm results in the same lift production as a 1 m long swallow-tail (Fig. 6)
Drag
The predominant influences on drag are similar to those on lift.
(1) Tail shape and length only really impact on drag at high airspeeds (Fig. 7)
(2) The Norberg effect can be seen to reduce drag particularly at high airspeeds for swallow tailed birds and generally reduces drag for all forked tailed birds (Fig. 7). At relatively large tail lengths as the Norberg effect reduces drag substantially it is theoretically possible for drag to become negative, this can be seen by comparing the drag produced at 14 ms-1 for a wren Troglodytes troglodytes (10 g) and a pheasant (Phasianus colchicus) (1.5 kg). The Norberg effect obviously decreases drag more for swallow tailed birds than for other forked tailed birds - this is reflected by the triangular shape of the area where the drag surface intersects the plane where drag is zero (Fig.7B).
(3) The impact of increasing tail area as all tail feathers elongate dominates the figure and quickly overwhelms the Norberg effect. This effect results in drag rapidly increasing with tail shape and length as tail shapes moves between -1 and -2 and between 1 and 2.
Power
Power is calculated for both level and turning flight. In turning flight the power surface has a similar shape as for the drag surface. Theoretically, power requirements can become negative when tails are sufficiently long and the Norberg effect operates (i.e. when tails are to some extent forked) (Fig. 8). When many tail feathers are elongated power requirements increase dramatically, especially at high airspeeds. The power required for level flight is symmetrical around a tail shape of zero as the Norberg effect does not operate when tails are furled (Fig. 8). The power required for level flight is also lower than that required for turning flight as the total drag is lower, again due to the furled tail.
Turn radius
Minimum turn radius was calculated for both gliding and flapping flight and these results are shown as two surfaces in Fig. 9. The upper surface is the minimum turn radius in flapping flight and the lower surface is the minimum turn radius in gliding flight. Turn radii at low speeds are large when tails are short but drop rapidly as tail increases for all tail shapes except for the region 0.2 to 1 (pintails to wedge-shaped tails). Turn radius drops for tails in which all feathers are elongated as a result of the increasing lift of the increasing tail area (turn radius drops with increasing lift). Turn radius also drops for all birds with forked tails (-0.2 to -1) as the Norberg effect increases lift, turn radii are smallest for swallow-tailed birds as here the Norberg effect has the greatest effect. At high speeds the most obvious effect is the Norberg mechanism that reduces turn radii for all forked tailed birds.
DISCUSSION
The impact of elongated tails on the four mechanical parameters considered here was very variable. The effect on body mass is negligible even when tails are large, changes in body mass are only likely to be important when small birds have very large tails. The turning moment of the tail is similar in that only large tails are likely to have significant effects on the static balance of the bird. However, the moment of inertia of the tail can rapidly become a significant component of the moment of inertia of the bird. This means that even relatively short tails can have large effects on the maximum rates of roll around the pitch and yaw axes of the bird. This will have similarly large effects on the agility of the bird unless the bird can increase the pitching and yawing forces it can produce. The moment of inertia of the tail may also be important, it is difficult to be precise about the significance of this variable as it is not known how a change in moment of inertia changes control. The muscles of the tail will have to generate force to overcome the moment of inertia of the tail. As the moment of inertia of the tail can increase by about 10 fold this will mean that the tail muscles will have to increase the force required to move the tail by a similar amount.
Mechanical effects will only be influenced by the size of the tail - both length and mass. The shape of the tail only has an impact if it changes the position of the centre of mass or mass of the tail. When aerodynamic effects are considered tail shape becomes important. Obviously the Norberg effect cannot operate unless tail streamers are present (i.e. the tail is forked) and the magnitude of the effect varies with the length of the tail streamers. Similarly the length and position of the maximum continuous span of the tail will be different in forked tails and wedge-shaped tails.
In general, the model predicts the familiar U- or J-shaped relations between flight parameters and airspeed. At low airspeeds lift, drag and power vary rather little with tail shape and length. However, at moderate to high airspeeds several clear patterns emerge between these flight parameters and tail shape and length. The general pattern to emerge is that the dominant effect on lift, drag and power is the impact of Thomas’s (1993a) delta wing theory. The delta wing effect only begins to operate as the size and/or position of the maximum continuous span of the tail changes. As the maximum continuous span increases lift also increases. As tail area in this region is also getting larger, drag increases and so does power. This delta wing effect is very large compared to the magnitude of the Norberg effect. However, the Norberg effect can be seen to increase lift, reduce drag and reduce power for forked tailed birds. The strong impact of the delta wing effect is likely to mean that tails in which all feathers are elongating are likely to evolve only in large species or in species which do not depend heavily on flight.
The exception to the general finding that the delta wing effect is stronger than the Norberg effect is when turn radii are considered. For minimum turn radius, the dominant effect is that of the Norberg mechanism. Turn radii are significantly smaller for all forked tailed birds than for any others. This would suggest that streamer tails are most likely to evolve in those species which depend on high levels of manoeuvrability - aerial hunting insectivores and raptors would be the most obvious groups.
The only available data of the magnitude of the Norberg effect are some measurements from wind tunnel experiments with model swallows (A.L.R. Thomas personal communication). These data were used in this model by incorporating a multiplication factor which operated in proportion to the size of the tail streamers relative to those of a swallow. This has produced negative drag and power in some situations when the length of the tail streamers is sufficient. This obviously means that the Norberg effect has been incorporated in this model incorrectly. In the absence of more information and data about the way in which this effect operates it is difficult to model the effect in any other way. At present it is even unclear whether it influences any birds other than swallows, although there seems no good reason to suppose that it does not operate on all streamertails as considered here.
In any species in which there are selective pressures for the evolution of an elongated tail all the factors considered here will impact upon the bird during the evolutionary process. However, it is clear that the evolutionary impact of each of these effects will be different for birds with different ecologies. A bird which only flies infrequently will obviously not incur the costs of flight with an elongated tail as much as a frequent flier. The costs of static balance may be important for such a species than the costs of increased drag. These considerations will have implications for the distributions of tail lengths and shapes across bird species. It is perhaps not surprising to find that a relatively terrestrial group like the pheasants (Phasianidae) have a large number of species in which tail extremely elongated.
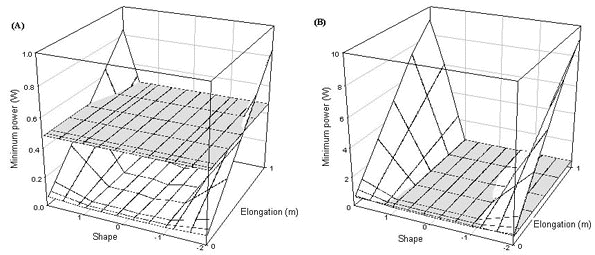
However, for any bird species which needs to fly for at least some of the time there are some thresholds on tail ornamentation due to the power needed to either fly at all or to fly efficiently. There are two crucial points in the on the power curve for flight - minimum power and maximum range. These are the minimum power required to fly and the power needed to fly at the speed that gives the greatest distance travelled per unit energy. If the power available to a bird does not exceed the minimum power then the bird will not be able to fly aerobically. If the power available does not exceed the maximum range power then it is unlikely to be capable of sustained flight. This will impact upon the ability of the bird to possess an ornamented tail if the power required to fly is increased through the possession of the elongated tail beyond the power available to the bird. This can be seen if the minimum power and maximum range power are calculated for a wren. All types of tail elongation increase the maximum range power beyond the power available and for only a small range of tail shapes is the minimum power for flight attainable (Fig. 10). A swallow on the other hand has a greater range of tail shapes available, only a few of the longest and largest tails take the minimum power requirements beyond the power available. Maximum range power is achievable for a swallow with all forked and wedge-shaped tails. Only tails in which all the feathers are elongated take the maximum range power beyond the power available (Fig. 11). It seems likely that at least some of the observed distribution of tail lengths and shapes is explicable because some tail designs are unavailable to some species.
It is possible to make some general statements regarding the distribution of tail shapes and lengths across species. From the arguments presented above it is clear that any form of tail elongation is unlikely to evolve in small species of birds. The first types of elongation to appear should be swallow-tails and then other forked tails (indeed there are strong arguments for shallow forks or swallow tails being the aerodynamically optimum tail design). Swallow tails should be particularly likely to evolve in species requiring high manoeuvrability. Tails in which all tail feathers are elongated should occur only in large species and in those species that have a low dependence on flight. Once all tail feathers are elongating there is rather little difference between elongated forked tails and elongated wedge-shaped tails. Conversely, the aerodynamics of a swallow-tail are very different from those of a pintail.
ACKNOWLEDGEMENTS
I would like to acknowledge the help of Adrian Thomas who wrote the initial version of the program used here - what seems like many years ago. Tim Benton was there when I got stuck. Chris Evans provided essential advice and solved some of the problems encountered while I was trying to find mathematical solutions to some aspects of this model, he also had the patience to check all my calculations.
REFERENCES
Aldridge, H.D.J.N. 1986. Manoeuverability and ecology in British bats. Myotis 23-24: 157-160.
Alexander, R.McN. 1983. Animal Mechanics; 2nd ed. Oxford: Blackwell Scientific Publications.
Andersson, M. 1994. Sexual selection.. Princeton; Princeton University Press: 597pp.
Balmford, A., Thomas, A.L.R. & Jones, I.L. 1993. Aerodynamics and the evolution of long tails in birds. Nature, London 361: 628-630.
Bilo, D., Bilo, A., Müller, M., Theis, B. & Wedekind, F. 1985. Neurophysiological-cybernetic correlates of course control in the pigeon. In: Nachtigall, W. (ed.) Biona Report 3, Bird flight - Vogelflug. Stuttgart; Gustav Fischer. 445 - 477.
Dudley, R. & Ellington, C.P. 1990. Mechanics of forward flight in bumblebees: II. quasi-steady state lift and power requirements. Journal of experimental Biology: 148, 53 - 88.
Evans, M.R. & Thomas, A.L.R. 1992 The aerodynamic and mechanical consequences of elongated tails in the scarlet tufted malachite sunbird: Measuring the cost of a handicap. Animal Behaviour 43: 337-347.
Hertzog, K. 1968. Anatomie und flugbiologie der vögel. Stuttgart; Gustav Fischer, Jena; V.E.B. Gustav Fischer.
Hughes, P.M. 1990. Aspects of microchiropteran reproduction with relevance to flight performance. PhD Thesis, University of Bristol, Bristol, U.K.
King, A.S. & King, D.Z. 1979. Avian morphology: general principles. In: King, A.S. & McLelland, J. (eds) Form and function in birds. New York; Academic Press. 1-38.
Lee, M. & Ho, C.M. 1990. Lift force of delta wings. Applied Mechanics Review 43, 9: 209-221.
Marden, J.H. 1994. From damselflies to pterosaurs: how burst and sustainable flight performance scale with size. American Journal of Physiology 266: 1077-1084.
Norberg, R.Å 1994. Swallow tail-streamer a mechanical device for self deflection of tail leading edge, enhancing aerodynamic efficiency and flight manoeuverability. Proceedings of the Royal Society of London B. 257: 227-223.
Pennycuick, C.J. 1975. Mechanics of flight. In: Farner, D.S. & King, J.R. (eds) Avian Biology. Vol. 5; New York; Academic Press: 1-75.
Pennycuick, C. J. 1989. Bird flight performance: a practical calculation manual. Oxford; Oxford University Press:
Rayner, J.M.V. 1979a. A vortex theory of animal flight. Part 1. The vortex wake of a hovering animal. Journal of Fluid Mechanics 91: 697 - 730.
Rayner, J.M.V. 1979b. A vortex theory of animal flight. Part 2. The forward flight of birds. Journal of Fluid Mechanics 91: 731 - 763.
Rayner, J.M.V. 1987. Form and function in avian flight. Current Ornithology 5: 1-66.
Thomas, A.L.R. 1993a. On the aerodynamics of birds' tails. Philosophical Transactions of the Royal Society of London B 340: 361-380.
Thomas, A.L.R. 1993b. The aerodynamic costs of asymmetry in the wings and tail of birds: asymmetric birds can't fly round tight corners. Proceedings of the Royal Society of London B. 254: 181-189.
Thomas, A.L.R. 1996. Why do birds have tails? The tail as a drag reducing flap, and trim control. Journal of Theoretical Biology 183: 247-253.
Fig. 1. Classification of tail shapes as used in the model.
Fig. 2. Tail mass as a percentage of total body mass for a wren.
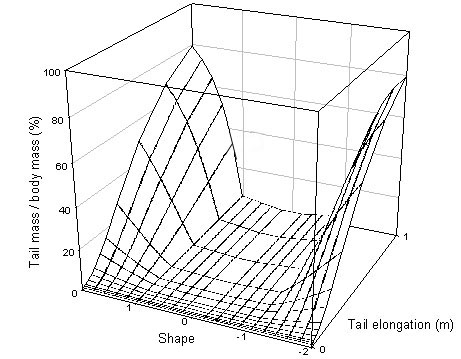
Fig. 3. Turning moment of the tail for a wren.
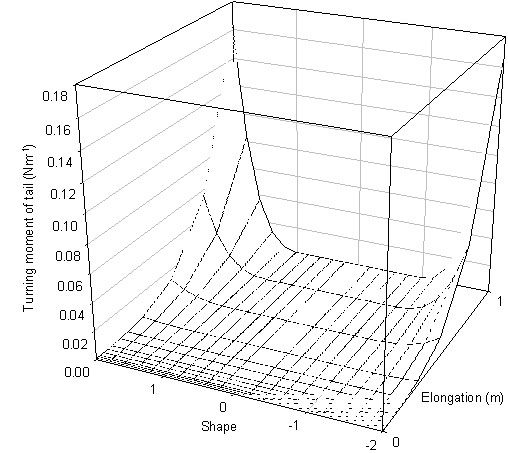
Fig. 4. Moment of inertia of the tail around the bird’s centre of gravity as a percentage of total moment of inertia for a wren.
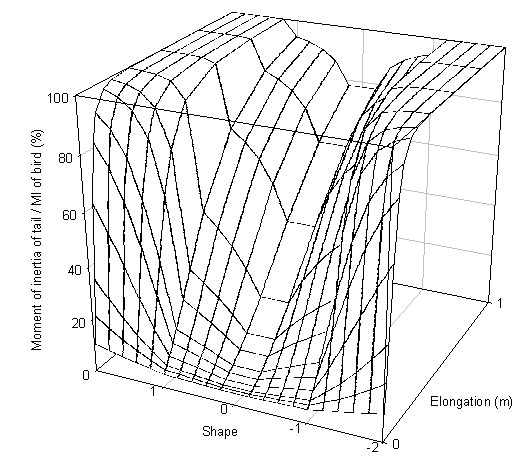
Fig. 5. Moment of inertia of the tail around tail attachment for a wren.
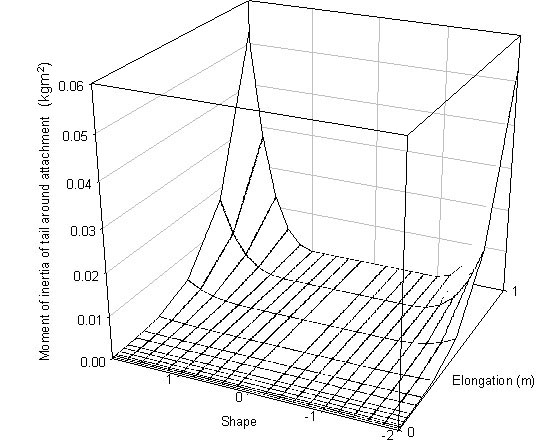
Fig. 6. Lift produced by a wren at (A) 5 ms-1 and (B) 14 ms-1.
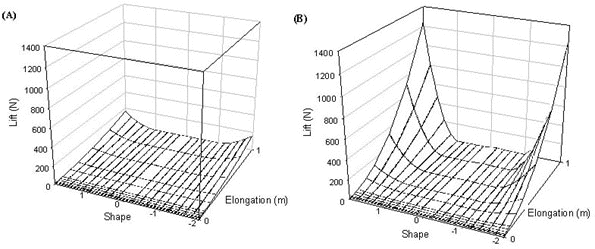
Fig. 7. Drag on a wren at (A) 5 ms-1 and (B) 14 ms-1 and (C) a pheasant at 14 ms-1.
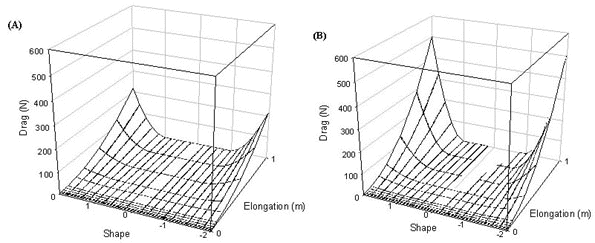
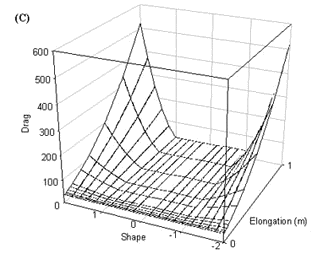
Fig. 8. Power required by a wren in turning flight at (A) 5 ms-1 and (B) 14 ms-1 and (C) in level flight at 14 ms-1.
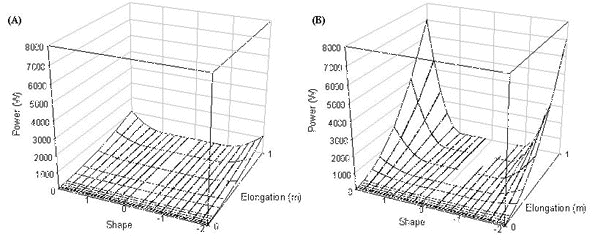
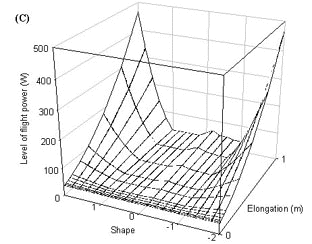
Fig. 9. Minimum turn radius of a pheasant at (A) 5 ms-1 and (B) 14 ms-1, the upper surface is for flapping flight the lower surface for gliding flight.
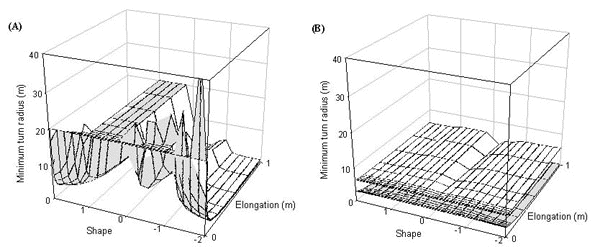
Fig. 10. Power required by a wren to fly at minimum power, the horizontal surface is an estimate of power available for flight (0.25W).
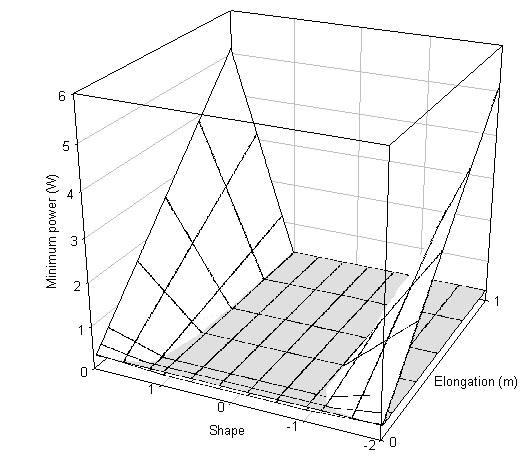
Fig. 11. Swallow (A) minimum power and (B) maximum range power, the horizontal surface is an estimate of power available for flight (0.47W).