S27.3: Is size important? Preliminary indications from the Birds in Reserves Project for the Woodland Biome
James A. Harrison & Rene A. Navarro
Avian Demography Unit, University of Cape Town, Rondebosch 7701, Cape Town, South Africa, fax 27 21 653726, e-mail batlas@maths.uct.ac.za
Harrison, J.A. & Navarro, R.A. 1999. Is size important? Preliminary implications from the Birds in Reserves Project for the Woodland Biome. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1538-1545. Johannesburg: BirdLife South Africa.South Africa, Lesotho and Swaziland have several hundred protected areas which vary greatly in size and are distributed unevenly in space. The role which these areas play in the conservation of biodiversity was analysed using the databases of the Southern African Bird Atlas Project (SABAP) and the Birds in Reserves Project (BIRP). Trends in the relative abundance of terrestrial species were analysed for areas inside protected areas (BIRP data) relative to other areas (SABAP data), and further analysed with respect to the size of protected areas and species guilds. The relative importance of different size classes of protected areas was evaluated and the guilds most dependent on these areas identified. The importance to certain species of relatively large protected areas, and of areas outside formally protected reserves, was demonstrated.
INTRODUCTION
South Africa, Lesotho and Swaziland have several hundred protected areas which vary greatly in size and are distributed unevenly in space. The role which these areas play in the conservation of biodiversity was analysed using the databases of the Southern African Bird Atlas Project (SABAP) and the Birds in Reserves Project (BIRP). Trends in the relative abundance of terrestrial species were analysed for areas inside protected areas (BIRP data) relative to other areas (SABAP data), and further analysed with respect to the size of protected areas and species guilds. The relative importance of different size classes of protected areas was evaluated and the guilds most dependent on these areas identified. The importance to certain species of relatively large protected areas, and of areas outside formally protected reserves, was demonstrated
In a country where land issues are at the top of the political agenda, and where conservationists are hard-pressed to acquire - indeed, even to keep - large areas for the protection of wildlife, the question 'How much land do we need?' is especially relevant. The Birds in Reserves Project (BIRP) focuses on the avifaunas of protected areas in order to answer this and other conservation-related questions (Harrison 1995).
Various studies have contributed to the SLOSS debate (Dobson 1995; Simberloff & Abele 1982), that is, is it better to have a Single Large protected area Or Several Small ones? Indications are generally that single large is better because it is able to preserve larger populations which are less likely to become extinct as a result of unpredictable events such as epidemics, fire, drought, etc. Large areas also encompass greater proportions of whole ecosystems, allowing for more effective management and preservation of ecosystem functioning. However, the Single Large option becomes less attractive when one has a diversity of habitat types and biotic communities which are spread over a wide area – too wide an area to encompass all within one reserve. The imperative to conserve biotic diversity then dictates that Several Small reserves need to be created so that a representative sample of all habitat types and communities are included in a network of protected areas. The question then becomes: 'How small dare we go?' In a South African context it also leads to questions like: 'Shouldn’t we sell off parts of the Kruger National Park to finance the purchase of land in the Little Karoo?' The question of minimum size for a wildlife refuge is critical because of the well-established fact that smaller fragments of habitat are more vulnerable to the loss of biodiversity than larger ones (Dobson 1995).
South Africa with its immense diversity of biomes and species (Siegfried 1989) faces this dilemma like few other countries. The issue is complicated by the fact that the answer to the question of minimum size will not be the same for different plants and animals. Nevertheless, perhaps we can begin to define the requirements for some taxa, including birds.
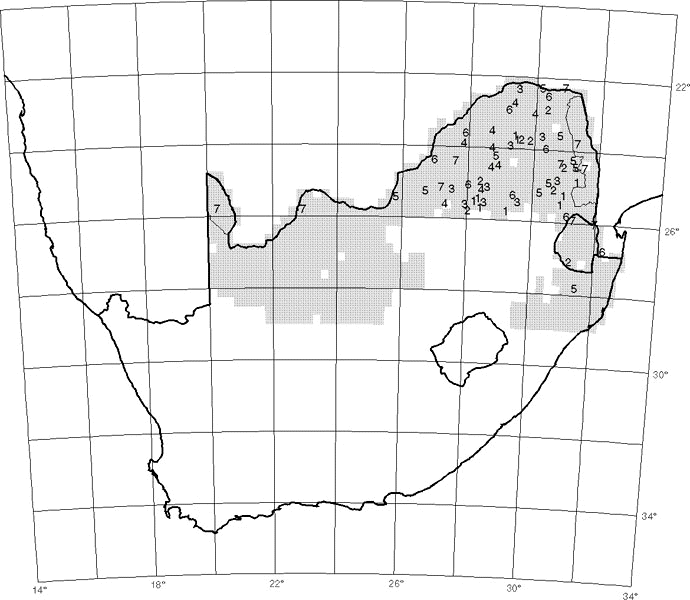
In South Africa there is an extensive area, and a large number of species, which comprise the woodland or savanna biome (Rutherford & Westfall 1986). This biome includes a large number of protected areas of widely varying size (Fig. 1). These provide an opportunity to investigate the occurrence of woodland birds in reserves of different sizes and thereby begin to describe the relevance of area to the preservation of different species.
METHODS
BIRP data consist of checklists of birds seen or heard during one day, or a minimum of three hours, within the boundaries of a defined protected area (PA) (Harrison 1995). The PAs used in this study are those located within the woodland biome, and having at least four BIRP checklists (4-3004, mean=92; excluding the Kruger National Park: 4-802, mean=45); these numbered 64 PAs (Fig. 1). Woodlands to the south of 29
°S are somewhat atypical of the biome as a whole, and were therefore not included.‘Woodland species’ used in the analyses were all terrestrial species with ranges falling wholly or substantially within the woodland biome. Many of these species also occur in other biomes, but species which are marginal to the woodland biome were excluded, as were all waterbirds. The number of woodland species, as defined, was 318.
A chi-square test was used to detect significant differences in the presence and absence of species in relation to the areas of the PAs. The data for each species were arranged in 2x7 contingency tables based on seven size classes of PA, and presence/absence. The size classes were: Class 1: <500 ha (n=9); Class 2: 500 to 2300 ha (n=8); Class 3: 2301 to 3000 ha (n=9); Class 4: 3001 to 5000 ha (n=9); Class 5: 5001 to 8000 ha (n=10); Class 6: 8001 to 20000 ha (n=9); and Class 7: >20000 ha (n=10). The limits of the classes were determined a priori such that the sample sizes were nearly equal, and do not have any significance in themselves.
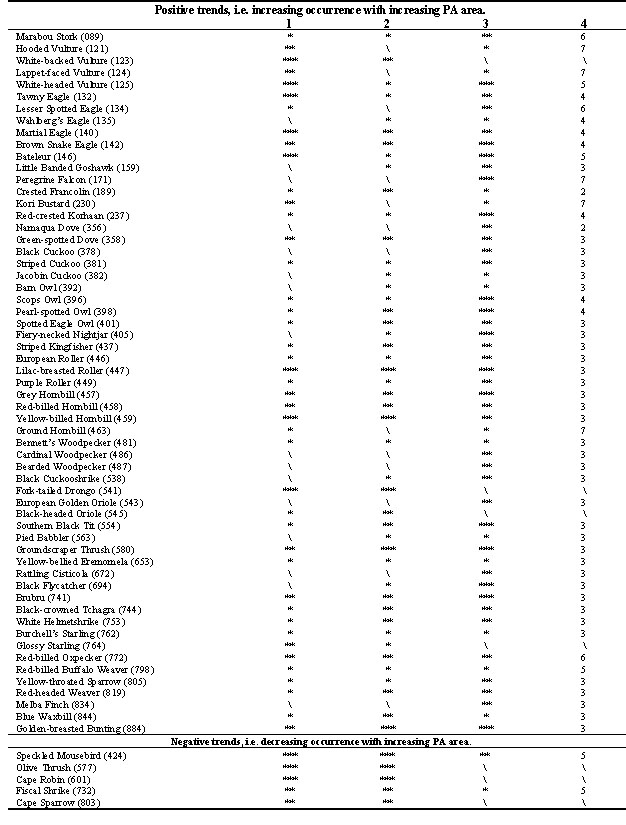
For those species with statistically significant results, the values of the residuals (deviations from the expected values) in the 2x7 contingency tables were scrutinised to determine at which size class the frequency of occurrence (i.e. the proportion of PAs in which the species was present) changed most, relative to the previous smaller class, by finding the largest change in the residuals. Such classes were termed ‘critical size classes’ (Table 1, column 4) and were related to the body sizes of the relevant species (Maclean 1993) by means of a test for linear trends in a contingency table (Sachs 1982) (Table 2).
A linear regression measured the strength of the relationship between reporting rate and log PA area. Reporting rate is the proportion of checklists which record a particular species; reporting rates have been used as an index of relative abundance to compare population densities in different parts of a species’ range (Harrison & Underhill 1997). The regression was done twice, firstly with the Kruger and Kalahari Gemsbok National Parks included, and secondly with them excluded. This was done because these two parks, although both within the woodland biome, are located in areas which are not represented by many other PAs, namely the lowveld and the Kalahari; also their very large size makes them atypical.
RESULTS
Because the analyses were based on presence/absence checklists, and not on direct measures of density, only relatively high levels of statistical significance were taken into account, i.e. the P<0.05 level of probability was not used, except in a few cases where the results of other tests strengthened the overall trend for the species. 64 species (20%) had results which indicated significant relationships between PA area and occurrence (Table 1). A further 75 (24%) had only weakly significant results (P<0.05), but were generally not considered further. 178 species (56%) showed no significant relationships.
Of the 64 species with significant results, 59 showed a positive relationship between occurrence and area of PAs, while 5 showed a negative relationship. The relationship between critical size class of PA and body mass, is highly significant (chi-square for linear trend = 30.8, 1 d. f., P=<0.001) (Table 2).
DISCUSSION AND CONCLUSIONS
Presence/absence data and reporting rates are crude measures of occurrence. Reporting rates are not good at discriminating between situations where a species is common but occurs at different levels of abundance. Conversely, ‘presence’ as defined for the chi-square test might represent only rare visits by a species to a locality. It is probable that many more species would have shown significant results had the quantitative data been based on actual measures of density. Despite the limitations of semi-quantitative data, the number of species for which clear trends emerged is striking.
The five species with negative trends in occurrence relative to PA size (Speckled Mousebird Colius striatus, Olive Thrush Turdus olivaceus, Cape Robin Cossypha caffra, Fiscal Shrike Lanius collaris, Cape Sparrow Passer melanurus) are informative in that they are all species with well-known associations with modified habitats in agricultural and suburban landscapes. Indeed some of them, e.g. the Cape Sparrow, probably occur within the biome only as a result of man-made changes to the landscape. Their higher reporting rates in smaller PAs could reflect a greater degree of ‘spill-over’ or immigration from areas outside of the PAs, that is, an edge effect. Alternatively it could reflect an ability to maintain populations in the absence of competitors. In either case, the trends for these species give us confidence in the positive trends for other species which are not generally associated with modified habitats.
A much larger number of species showed positive trends in occurrence relative to PA size. In this group it is striking that the species represent a wide range of body sizes, feeding guilds and taxonomic groups, but raptors (including owls and vultures) and hornbills and, to a lesser extent, woodpeckers, rollers, shrikes and cuckoos, all appear to be particularly well represented. Predators on vertebrates can be expected to be dependent on relatively undisturbed wildlands to provide sufficient density and diversity of prey, unless they are rodent specialists. Hornbills, rollers and shrikes, also depend partly on vertebrate prey (Maclean 1993). Several raptors are directly or indirectly persecuted by humans (Oatley et al. 1998), particularly on farmlands, making large reserves a vital refuge for these species (e.g. Simmons 1997) because they allow the birds to carry out all their activities while remaining within the boundaries of the refuge at all times.
Woodpeckers use dead wood extensively as a source of invertebrate prey; they and several other species are also dependent on holes in dead trees for nesting sites (e.g. du Plessis 1997). Dead wood is used as fuel by rural communities and is therefore a scarce resource outside of protected areas. In many cases we simply know too little about the relative importance of different prey species, and it may be that the caterpillars preferred by a particular species of cuckoo, for example, are simply not available outside of protected areas where plant communities are still intact.
Although these results are preliminary, it does appear that we can draw the conclusion that many woodland species occur more often in larger reserves than in smaller ones and, in addition, may occur at higher densities in larger protected areas. Furthermore, the analysis of critical size classes shows a clear relationship between body size and PA area, something which could have been predicted on the basis of relative territory sizes. It is striking how large a proportion of the species with positive trends (n=35, 65%) have Class 3 (2301 to 3000 ha) as their critical size class. This suggests that an area of the order of 2500 ha may be an effective minimum for many small- to medium-sized woodland species, while considerably larger areas are needed for species of large body size. Notably, four large species (Hooded Vulture Necrosyrtes monachus, Lappet-faced Vulture Torgos tracheliotos, Kori Bustard Ardeotis kori and Ground Hornbill Bucorvus leadbeateri) have Class 7 (>20000 ha) as their critical size class. As suggestive as these critical size classes are, no firm conclusions can be drawn without further investigation.
It is well known that species richness increases with land area. It can be argued that the positive trends shown here are merely an artifact of the inclusion of a greater diversity of woodland and other habitats as area increases, having little if anything to do with area per se (Connor & McCoy 1979). Our response is that this is unlikely to be true for large species, such as vultures, with large foraging ranges, but may well be true for smaller species. Nevertheless, even if habitat diversity is the dominant factor determining richer communities of woodland birds, it remains relevant that larger areas tend to include greater habitat diversity, and consequently a greater diversity of woodland species. It is quite probable that many woodland birds depend on a complex mix of habitats, even if they spend most of their time in only one type. The needs and behaviours of birds, even resident species, also tend to vary seasonally, and with irregular events such as fire, drought, and flood.
On the other hand, factors operating from outside of the protected area, particularly the condition and linkage of neighbouring unprotected lands, will tend to be more influential on small refuges than larger ones because of their relatively high edge-to-area ratios. Many of the smallest reserves in the study area are those most embedded in urban and peri-urban landscapes and therefore largely isolated from other similar habitat patches. The value of relatively large protected areas is probably even greater than indicated by this study because many refuges in rural contexts benefit from linkage to extensive areas of privately owned woodlands on cattle and game farms which provide corridors and ‘stepping stones’ between protected areas. If these privately owned lands were to become degraded, effectively isolating the protected areas, more species’ ability to survive would be negatively affected (e.g. Recher & Serventy 1991; Soulé et al. 1988).
The fact that the presence and abundance of so many species of woodland birds, as mobile as they are, show a positive correlation with the size of protected areas, is pertinent to conservation planning. The clear need to establish several refuges in a network of protected areas which is representative of the total biodiversity, together with the demonstrated inability of small reserves to maintain their initial species richness (e.g. Soulé et al. 1988), creates a real tension in the formulation of conservation policy. The problem is exacerbated by financial constraints to the establishment or enlargement of reserves. What this study shows is that the size of refuges in the woodland biome is relevant even to such mobile species as birds, and that the smaller reserves are not effective in preserving the full diversity of woodland birds. Amongst the implications of this finding are a need for clearly defined conservation objectives for any particular refuge, and the necessary aut-ecological information and management strategies to support those objectives.
ACKNOWLEDGEMENTS
Sponsorship of the Birds in Reserves Project by The Tony and Lisette Lewis Foundation South Africa, is gratefully acknowledged.
REFERENCES
Connor, F.C. & McCoy, E.D. 1979. The statistics and biology of the species-area relationship. The American Naturalist 113: 791-833.
Dobson, A.P. 1996. Conservation and biodiversity. New York; Scientific American Library.
Du Plessis, M.A. Redbilled Woodhoopoe Phoeniculus purpureus. In: Harrison, J.A., Allan, D.G., Underhill, L.G., Herremans, M., Tree, A.J., Parker, V. & Brown, C.J. (eds) The atlas of southern African birds. Vol. 1 Non-passerines. Johannesburg; BirdLife South Africa: 202-203.
Harrison, J.A. 1995. Birds in Reserves Project: instructions. ADU Guide No. 1. Cape Town; Avian Demography Unit.
Harrison, J.A. & Underhill, L.G. 1997. Introduction and methods. In: Harrison, J.A., Allan, D.G., Underhill, L.G., Herremans, M., Tree, A.J., Parker, V. & Brown, C.J. (eds) The atlas of southern African birds. Vol. 1 Non-passerines. Johannesburg; BirdLife South Africa: xlv-lxiv.
Maclean, G.L. 1993. Roberts’ birds of southern Africa. Cape Town; John Voelcker Bird Book Fund.
Oatley, T.B., Oschadleus, H.D., Navarro, R.A. & Underhill, L.G. 1998. Review of ring recoveries of birds of prey in southern Africa: 1948-1998. Johannesburg; Endangered Wildlife Trust.
Sachs, L. 1982. Applied statistics: a handbook of techniques. New York; Springer-Verlag.
Recher, H.F. & Serventy, D.L. 1991. Long term changes in the relative abundances of birds in Kings Park, Perth, Western Australia. Conservation Biology 5: 90-102.
Rutherford, M.C. & Westfall, R.H. 1986. Biomes of southern Africa – an objective categorization. Memoirs of the Botanical Survey of South Africa 54: 1-98.
Siegfried, W.R. 1989. Preservation of species in southern African nature reserves. In: Huntley, B.J. (ed.) Biotic diversity in southern Africa: concepts and conservation. Cape Town; Oxford University Press: 186-201
Simberloff, D. & Abele, L.G. 1982. Refuge design and island biogeographic theory: effects of fragmentation. The American Naturalist 120: 41-50.
Simmons, R.E. 1997. Bateleur Terathopius ecaudatus. In: Harrison, J.A., Allan, D.G., Underhill, L.G., Herremans, M., Tree, A.J., Parker, V. & Brown, C.J. (eds) The atlas of southern African birds. Vol. 1 Non-passerines. Johannesburg; BirdLife South Africa: 202-203.
Soulé, M.E., Bolger, D.T., Alberts, A.C., Wright, J., Sorice, M. & Hill, S. 1988. Reconstructed dynamics of rapid extinctions of chaparral-requiring birds in urban habitat islands. Conservation Biology 2: 75-92.
Table 1. Statistically significant results of the regression analysis, including (column 1) and excluding (column 2) Kruger and Kalahari Gemsbok National Parks, and the chi-square analysis (column 3). The symbols *, **, and *** respectively denote the p<0.05, p<0.01 and p<0.001 levels of significance, and \ denotes no statistical significance. Critical size classes are given in column 4. The common names of species, and their numbers, are those used in Maclean (1993).
Table 2. Body weight of species (indicated by their Roberts’ numbers, as in Table 1) in relation to critical PA areas (see Table 1).
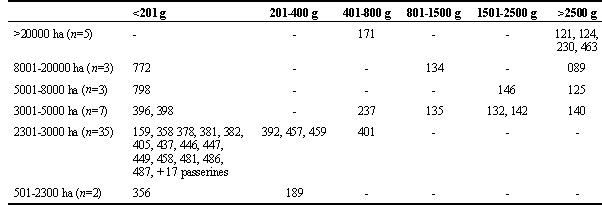
Fig. 1. Map of South Africa showing the extent of the woodland biome north of 29°S (shaded area), the boundaries of the Kruger National Park (in the north-east) and the Kalahari Gemsbok National Park (in the north-west) and the approximate centre-points of 62 other protected areas in seven size classes.