S24.5: Is the timing of breeding in Great and Coal Tits adapted to maximum nestling food or to avoid juvenile predation?
Beat Naef-Daenzer, Fritz Widmer & Maria Nuber
Swiss Ornithological Institute, CH-6204 Sempach, Switzerland, fax 41 41 462 9710, e-mail naefb@orninst.ch
Naef-Daenzer, B., Widmer, F. & Nuber, M. 1999. Is the timing of breeding in Great and Coal Tits adapted to maximum nestling food or to avoid juvenile predation? In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1406-1415. Johannesburg: BirdLife South Africa.Traits such as date, age or condition at which juvenile altricial birds leave the nest may have a great impact on survival and therefore fitness. In Great and Coal Tits Parus major, P. ater the physical condition of the chicks is crucial for their subsequent survival. Condition, however, is determined by the availability of food and the foraging performance of parents during the nestling period. Therefore, the parent's decisions on place and timing of breeding are important for both nestling growth and post-fledging survival. Actual knowledge, however, is restricted to the nestling period because visual recording of small birds outside the nest in dense vegetation is almost impossible.
Using new radiotracking tools, the present study provides evidence that post-fledging mortality is a major determinant of total reproductive success in both species. Juvenile mortality during the first three weeks after fledging was up to 54%. Post-fledging mortality was inversely correlated with fledging weight and increased during the season. Fledgling mortality causes considerable selection for parents having the chicks fledged as early as possible. Very early breeders profit from a minimal post-fledging mortality, but are constrained by low food availability. On the other hand broods that coincide with the maximum prey availability can fledge a large number of chicks but suffer from high post-fledging mortality. There is probably a trade-off between maximising nestling growth on one hand and minimising post-fledging mortality on the other hand. Evolutionary response to these counteracting selection processes is supposed to restrict the bird's breeding to a very short period.
INTRODUCTION
Keeping the offspring at a place where the young are protected against bad weather and predation is advantageous, most obviously in hole-breeding birds such as Great and Coal Tits Parus major, P. ater. This strategy, however, involves considerable parental costs. Large amounts of food have to be carried to the nest before the young will be able to visit feeding sites on their own. As a consequence, the timing of breeding in relation to food availability is critical because tits have large broods, and caterpillars (which make up to 85% of the diet) are available for a short period only (e.g. Perrins 1992). Thus, pairs that have their brood coincide with the peak availability of caterpillars contribute more to the breeding population than those breeding too early or too late (Tinbergen & Boerlijst 1990, Perrins 1992, Van Noordwijk et al. 1995, Verboven & Visser 1998). With regard to the nestling period, the availability of food is considered the main environmental determinant of nestling growth and condition causing selection that restricts the tit's breeding onto the period of maximum prey availability (Tinbergen & Boerlijst 1990, Van Noordwijk et al. 1995, Verboven & Visser 1998). This selection, however, is not likely to occur during the nestling period itself because nestling mortality is low (5-10% in first, 15-25% in second broods (Van Balen 1973, Glutz 1993)) compared with the survival from fledging until the following breeding season of only 5-30% (Tinbergen & Boerlijst 1990, Drent 1984). Energetic constraints and predation have been supposed to be factors affecting juvenile survival (Van Balen 1973, Drent 1984, Perrins 1980).
A methodological bottleneck in assessing the fitness consequences of variation in the timing of breeding and other reproductive traits is the estimation of the post-fledging survival in different cohorts of juvenile birds. Recruitment rates (i.e. the number of birds present in the breeding population of the subsequent year) have been shown to vary with important reproductive traits. However, this estimate is affected by various factors such as mortality during different seasons which in turn is affected by predation, food availability or social mechanisms. Additionally, estimates may be biased by dispersal out of the study area. The interference of all these factors is likely to confound the effect of the target variables on reproductive traits. To solve the problem, monitoring the juvenile birds beyond the nestling phase is indispensable. However, enormous technical problems to follow juveniles in dense vegetation prevented access to this part of the biological system.
In this radiotracking study we present an approach to assess the post-fledging ecology and mortality of small woodland birds. We analyse the survival of juvenile tits during the post-fledging period and investigate factors that determine their survival during the first three weeks after fledging. On this basis we quantify to what extent post-fledging mortality is related to the parents' timing of breeding and therefore its relevance for evolutionary response to environmental conditions.
METHODS
We developed new radio-tracking tools allowing birds of a minimal body mass of 7g to be tracked. Because a radio-tag must not affect the animal's health and behaviour, the radiotracking of bird fledglings (of 8-11 g in the Coal Tit and 14-19 g in the Great Tit, respectively) requires very small transmitters and extremely careful treatment. The new transmitter is a simple self-pulsing oscillator design which operates with a minimum of electronic components (Cochran 1963, Kenward 1978, Naef-Daenzer 1984). The transmitter without battery is 6x6x1.5 mm and has a mass of 170 mg. Zinc-Air cells of 290 mg lasting for 20-30 days were used. The total mass of the tag therefore was 460 mg, which is 2.4-3.3% of a Great Tit and 4.2-5.8% of a Coal Tit fledgling, respectively. The investigation was undertaken in a mixed deciduous forest on the 'Blauen' hill, at 300-600 m a.s.l. near Basel, NW-Switzerland. From 1995 to 1997 we tagged a total of 221 fledglings. The tags were either glued into the feathers of the back (using 20-40 mg of cyanoacrylate, n = 31, Raim 1978) or mounted using a Rappole-harness (c. 18 mg, n = 190, Rappole & Tipton 1990) made from fine cotton string. To prevent premature fledging, tags were mounted either three to four days before fledging or immediately before leaving the nest. A control group of 121 nestlings was individually colour-marked using waterproof pencils or left unmarked (1 per brood). Birds of less than 8 g (Coal Tit) or 15 g (Great Tit) were colour-marked but not equipped with transmitters. In order to establish maximal security against transmitter losses and failures, three to four chicks of a brood were radio-tagged and the remaining birds were colour-marked. The median rate of technical transmitter failures (excluding loss for improper mounting) was 1.8% per day. All individuals of a family can be recorded permanently as long as one transmitter is functioning. In some broods one of the adult birds also carried a transmitter.
Transmitters were first tested on captive juvenile tits. We noticed no differences in behaviour and manoeuvrability of tagged and untagged birds. With regard to mortality, a quantitative control of possible transmitter effects is possible and reveals no effect (Fig. 1).
During the first 20 days after fledging, the families remain together and no juvenile dispersal was observed. Death of juveniles was recorded when predation was observed visually or at the receiver (i.e. rapid movements out of the family group with an abrupt break of the signal), when dead birds or parts were found, or when birds were not observed for 48 hours.
RESULTS
A total of 3495 bird-days were recorded during which 162 birds (47%) died. There was no difference in the mortality rates of colour-marked and radio-tagged birds (ANCOVA, F1;83 = 0.045, P>0.8). We therefore conclude that the mortality data are not biased by the transmitter load. Furthermore, this result shows that there was no significant overestimation of mortality because of either unrecognised transmitter failure or reduced observation probability of colour-marked birds. Otherwise the curves would differ in intercept and/or slope. For the period of 20 days after fledging there was no significant difference in the mortality of the two species (ANCOVA, F2;83 = 0.17, P>0.68). Thus, the data from both species were pooled for most of the further analyses.
None of the 115 radio-tagged juveniles surviving for more than two weeks after fledging was observed to disperse out of the study area. This indicates that the birds disappeared from the family groups had died. In most cases, however, mortality was neither observed directly nor were the carcasses found (see methods). The 26 cases of direct observations or retrieval of carcasses indicate that the major cause of fledgling mortality was predation by Jays Garrulus glandarius, Great Spotted Woodpeckers Dendrocopos major, Sparrowhawks Accipiter nisus and martens Martes sp. A small proportion of juvenile birds was found starved or dead from unknown causes (14 out of the 221 radio-tagged individuals, untagged animals cannot be found when dead). The high frequency of observations of potential predators visiting the family groups also suggests that predation was one major cause of mortality in the post-fledging period although it is not possible to quantify to what extent.
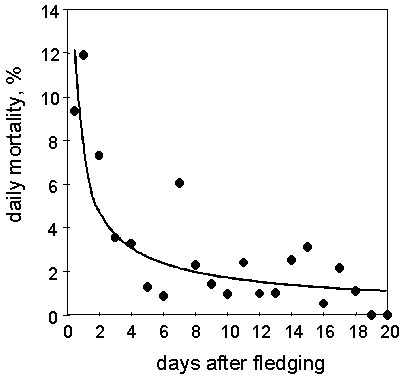
The number of surviving juveniles in relation to the time after fledging follows logarithmic function (y = 81,92-21.4·logx, r = -0.98). Consequently, the daily mortality rate decreases in a potential function (y= 7.60.x-0.65, n = 21 days, r = -0.74, P<0.001 (Fig. 1). This result demonstrates that there is a very short period of high mortality. 32% of the juveniles were lost during the first 4 days after fledging, but thereafter the daily mortality quickly decreases and levels off. From day 15 onwards, the average daily mortality rate had dropped to 1.1%. This is still high in relation to the adult mortality of 0.15% per day (or 50% p.a.).
The body condition of the fledglings strongly affects their survival after fledging. Fig. 2 gives the proportion of surviving great and Coal Tits in relation to their fledging weight. The data of each species were split into two cohorts with fledging weight above and below the three-year average, respectively. In both species, low fledging weight resulted in a markedly reduced survival. In the Great Tit, however, the difference in mortality was much more pronounced than in the Coal Tit (ANCOVA, F1;82 = 26.84, P<0.0001 for difference in slope, F = 4.25, P<0.04 for difference between species). None of the juveniles with very low fledging weights (0.6-0.8 the average fledging weight, n = 22) survived longer than 3 days after fledging. Since fledging weight is considered an indicator of physical condition these large differences in mortality indicate that reduced growth during the nestling period greatly reduces the survival of juvenile tits during the post-fledging period. There was no significant difference in the average fledging weight of early and late birds, thus the result is not confounded by fledgling condition. (Great Tit: early 16.40 g ± 1.6(S.D.) n = 168, late 16.53 g ±1.81 n = 91; Coal Tit: early 10.70 g ± 0.96 n = 60, late 10.16 g ± 0.89 n = 13).
We found that the post-fledging survival shows large seasonal variation. In all three years, mortality rates were much higher in the late than in the early season. Of the birds fledged before June 15, 64 (33%) of 192 juveniles were lost within 15 days after fledging. By contrast, 89 (59%) of 151 birds fledged after June 15 died within the same period. (ANCOVA, interactions, F1,38 = 32.53, P<0.0001 (Fig. 3). The average fledging mass in the two periods was not significantly different in both species, indicating that the result is not confounded by fledging mass (Great Tit, early: 16.4 g ±1.6 (S.D.), late: 16.53 g ±1.81; Coal Tit, early: 10.7 g ±0.96, late: 10.16 g ±0.89). This large seasonal difference in fledgling mortality suggests that there is a great effect of the timing on the contribution of 'early' or 'late' birds to the breeding population in the next breeding season.
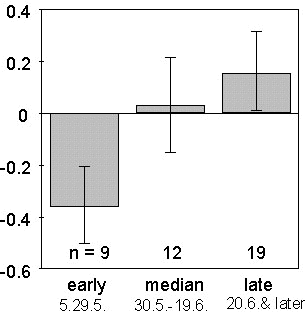
The seasonal changes in fledgling mortality result in markedly higher losses in the late broods. The most important question is whether these effects matter in relation to the whole population of fledglings. Based on our data on the fledglings produced per 5-day period and the observed mortality rates, we calculated selection differentials for the first three post-fledging weeks. The selection differential is the difference between the mean yearly survival and the observed survival in a given 5-day period weighted for the overall survival. Fig. 4 gives the average selection differentials for early, intermediate and late fledglings (early: before May 15, intermediate: between May 15 and June 14, late: after June 15). There was a strong increase of the selection differentials during the season (ANOVA, F2;37 = 31.147, P<0.0001). This signifies that offspring of early broods increased in proportion in the population of juveniles surviving the observation period, whereas the proportion of late fledglings was reduced.
DISCUSSION
Almost all of the fitness-relevant interactions between the breeding tits and their environment have so far been attributed to the laying and nestling period (e.g. Tinbergen & Boerlijst 1990, Van Balen 1973, Van Noordwijk et al. 1995). Our results demonstrate that post-fledging mortality greatly affects the reproductive success and that the parents' decisions on the timing of breeding have an impact far beyond the nestling period. Selection occurring during the post-fledging phase is a considerable factor for optimal timing of breeding and the evolution of the tit's reproductive system. Juvenile mortality in this phase is not a random process with a constant 'decay' rate (like radioactive decay). There is a short period of high vulnerability, great seasonal variance and a strong relationship between mortality and the physical condition of the fledglings. Given the presence of heritable variability in the reproductive traits there is much scope for evolutionary response to minimise the post-fledging loss of juveniles (Noordwijk et al. 1988, Gebhardt-Henrich & Van Noordwijk 1991, Keller & Van Noordwijk 1993).
It appears that the major proportion of the juvenile mortality from fledging till the next breeding season can be attributed to the very first weeks after leaving the nest. This gives a proximate explanation of the close correlation of recruitment rates with habitat conditions during the laying and nestling period such as temperature or development of prey populations. However, this does not imply that the causes of mortality and selection are related to the nestling period, too. We detected two main factors, fledging weight and date, both of them causing differentials in the survival of different cohorts of birds.
First, post-fledging survival strongly depends on fledging weight. This result confirms earlier reports (e.g. Tinbergen & Boerlijst, Verboven & Visser 1998) and underlines the importance of having the nestling period coincide with the maximum prey availability. The fledging weight results from growth during the nestling period and is therefore determined by the rate at which energy (in form of caterpillars) is delivered to the nest. The energy flow to the nest, however, is a correlate of density and sise of prey in the habitat, whereas the parents have little options to adjust their feeding rates if the brood requires more food (Perrins 1992, Naef-Daenzer & Keller, Nur 1984, 1986). Since starvation was observed rarely and the largest proportion of juveniles has apparently been predated, there is need to explain how predation rates correlate with fledgling condition. One possible mechanism is that birds of lower condition may beg more intensively and move more frequently which makes it easier for predators to locate and catch those birds in the dense canopy.
A contrasting relationship is the result that post-fledging survival declines with fledging date. This again confirms earlier analyses of recruitment rates (Kluyver 1951, Perrins 1965, 1970), indicating that juveniles from the earliest broods survive better even if they do not profit from the best feeding conditions. Since the major proportion of tit fledglings has been predated, this trend was probably caused by the increasing food requirements of the predators: For one, the Sparrowhawk Accipiter gentilis, several studies report observations that support this interpretation (e.g. Geer 1981, 1982, Newton & Marquiss 1982, Moss 1979). Similar to the tit-caterpillar system, Sparrowhawks adjust their breeding to the appearance of profitable and abundant prey: songbird fledglings. Newton & Marquiss (1982) have demonstrated that fledglings make up to 60% of the nestling diet in Sparrowhawks and that their breeding success seems to be causally linked to the availability of this prey. The proportion of titmice delivered to Sparrowhawk nests increased from mid May to end of June, which supports the interpretation that the date effect in post-fledging mortality was caused by increasing predation (Newton 1978). Furthermore, Geer (1982) provided evidence that the most vulnerable individuals i.e. the recently fledged young were caught at higher rates than the more experienced ones. This is in accordance with our finding that the maximum mortality rates were observed in the first days after fledging (Fig. 1).
Our (preliminary) results make clear that much of the post-fledging mortality (or survival) is determined by the timing of breeding which is a decision of the parents. It is well established that in order to maximise nestling growth tits adjust their nestling period to the maximum availability of prey. This, however, seems to be only one part of the system.
The finding that tits also must adjust their breeding in relation to the reproduction (i.e. the maximum food requirements) of their predators, is new and implies a trade-off between the energetic benefits of raising a brood when foraging conditions are best and the costs of increased post-fledging mortality: Nestlings that profit from the maximum food availability in the last week of May will fledge mid June and therefore suffer from a considerable fledgling mortality (Fig. 3 & Fig. 4). Our radiotracking results reveal that the timing of breeding in respect to post-fledging mortality is as important as it is to have the nestling period coincide with the peak abundance of food. Further research should develop these findings towards a model of how titmice parents deal with the two tasks: catching caterpillars optimally but avoiding that juveniles will be caught themselves.
ACKNOWLEDGEMENTS
We greatfully acknowledge the contribution of all assistants in the field: Ch. Angst, F. Buner, J. Dietrich, G. Gaidon, M. Grüebler, D. Hegglin, S. Joss, M. Keiser, D. Mülli, M. Neecke, R. Schwilch, U. Sieber, P. Steck, C. Steck, M. Winzer. Microbonding SA, CH-2023 Gorgier designed and pre-mounted the miniaturised prints. Quarz AG, CH-8617 Mönchaltorf provided technical support. The investigation was supported by the Swiss National Science Foundation.
REFERENCES
Cochran, W.W. & Lord. R.D. 1963. A radiotracking system for wild animals. J. Wildl. Manage. 27: 9-24.
Clobert, J., Perrins, C.M., McCleery, R.H. & Gosler, A.G. 1988. Survival rate in the Great Tit Parus major in relation to sex, age and immigration status. J. Anim. Ecol. 57: 287-306.
Drent, P.J. 1984. Mortality and dispersal in summer and its consequences for the density of Great Tits (Parus major) at the onset of autumn. Ardea 72: 127-162.
Gebhadt-Henrich, S. & Van Noordwijk, A.J. 1991. Nestling growth in the Great Tit. I. Heritability estimates under different environmental conditions. J. Evol. Biol. 3: 341-362.
Geer, T. 1981. Factors affecting the delivery of prey to nestling Sparrowhawks (Accipiter nisus). J. Zool. Lond. 195: 71-80.
Glutz, U.N. 1993. Handbuch der Vögel Mitteleuropas. Bd. 13(4). Wiesbaden.
Keller, L.F. & Van Noordwijk, A.J. 1993. A method to isolate environmental effects on nestling growth, illustrated with examples from the Great Tit (Parus major). Functional Ecology 7: 493-502.
Keller, L.F. & Van Noordwijk, A.J. Effects of local environmental conditions on nestling growth in the Great Tit Parus major L. Ardea 82:349-362.
Kenward, R. 1987.Wildlife radio tagging. Academic Press. London.
Naef-Daenzer, B. 1993. A new transmitter for small animals and enhanced methods of home-range analysis. J. Wildl. Manage. 57(4): 680-689.
Newton, I. 1978. Feeding and development of Sparrowhawk (Accipiter nisus) nestlings. J. Zool. Lond. 184: 465-488.
Newton, I. & Marquiss, M. 1982. Food, predation and breeding season in Sparrowhawks (Accipiter nisus). J. Zool. Lond. 197: 221-240.
Van Noordwijk, A.J., van Balen, J.H. & Scharloo, W. 1981. Genetic variation in the timing of reproduction in the Great Tit. Oecologia 49:158-166.
Van Noordwijk, A.J., van Balen, J.H. & Scharloo, W. 1988. Heritability of body sise in a natural population of Great Tits and its relation to age and environmental conditions during growth. Genet. Res. Camb. 51: 149-162.
Van Noordwijk, A.J., McCleery, R.H. & Perrins, C.M. 1995. Selection for the timing of Great Tit breeding in relation to caterpillar growth and temperature. J. Anim. Ecol. 64: 451-458.
Moss, D. 1979. Growth of nestling sparrowhawks (Accipiter nisus). J. Zool. Lond. 187: 297-314.
Perrins, C.M. 1980.Survival of young Great Tits. Acta XVII Congr. Int. Orn. Berlin.
Perrins, C.M. 1992. Tits and their caterpillar food supply. Ibis 133 suppl. 1: 49-54.
Raim, A. 1978. A radio transmitter attachment for small passerine birds. Bird-Banding 494: 327-332.
Rappole, J.H. & A. Tipton. 1990. New harness design for attachment of radio transmitters to small passerines. J. Field. Orn. 62(3):335-337.
Ricklefs. R.E. 1983. Avian postnatal development. In: Farmer, D.S. & J.R. King (eds.): Avian biology, Vol.7. Academic Press. London.
Tinbergen, J. M. & C.M. Boerlijst. 1990. Nestling weight and survival in individual great tits (Parus major). J. Anim. Ecol. 59:1113-1127.
Van Balen, H. 1973. A comparative study of the breeding ecology of the Great Tit (Parus major) in different habitats. Ardea 61: 1-93.
Verboven, N, & M. Visser. 1998. Seasonal variation in local recruitment of great tits: the importance of being early. Oikos 81(3): 511-615.
Fig. 1. Daily mortality rate of radio-tagged and colour-marked Great and Coal Tit fledglings in relation to time after fledging (y=7.60·x-0.651, n = 21 days, r=-0.74, P<0.001). Fledglings are most vulnerable during the first days after leaving the nest.
Fig. 2. Survival of Great and Coal Tit fledglings in relation to time and fledging weight. Percentages of surviving young are given for the cohorts of individuals above or below the three-year average fledging weight, respectively. In both species, poor physical condition results in increased post-fledging mortality (ANCOVA, F1;83 = 26.84, P>0.0001). In the Great Tit the variance in mortality caused due to fledgling condition is greater than in the Coal Tits (ANCOVA, interaction: F1;83 = 10.04, P<0.003). Circles: 143 Great Tits: y = 89.23-16.0x, r = -0.98, n = 21. Dots: 123 Great Tits: y = 75.02-22.72x, r = -0.99, n = 21. Open squares: 40 Coal Tits: y = 84.11-17.30x, r = -0.95, n = 21. Filled squares: 36 Coal Tits: y = 80.57-21.64x, r = -0.95, n = 21.
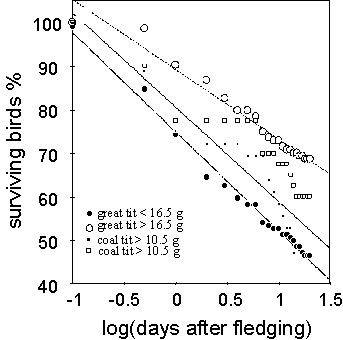
Fig. 3. Variation in mortality of Great and Coal Tits fledglings in relation to fledging date. Percentages of surviving young are given for the cohorts of individuals fledged before (dots) and after (circles) June 15. The mortality rate during the first 20 days after fledging was significantly higher in juveniles fledged late in the season (ANCOVA, df = 1;38, F = 32.53, P<0.0001).
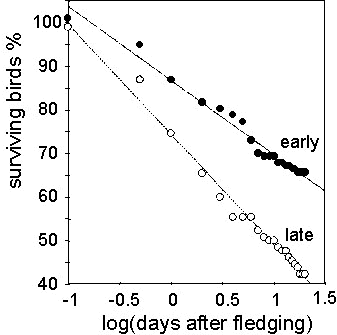
Fig. 4. Selection differentials for fledging date in relation to the date of fledging. The selection differential is the residual mortality of the cohort of birds fledged in a given 5-day period on the average yearly mortality weighted for the 3-year average. Averages and standard deviations are given for early, median and late broods, respectively. During the 20-day observation period, the proportion of early fledglings in the population of survivors increased, whereas the proportion of late fledglings declined, which signifies selection for early breeding (ANOVA, F2;37 = 31.15, P<0.0001).