S24.4: Post-fledging behaviour, dispersal and survival in Eurasian Sparrowhawks
Ian Newton1, Peter Rothery2 & Ron Frumkin3
1Institute of Terrestrial Ecology, Monks Wood, Abbots Ripton, Huntingdon, Cambridgeshire PE17 2LS, UK, fax 44 0 1487 773467, e-mail i.newton@ite.ac.uk; 2Institute of Terrestrial Ecology, Monks Wood, Abbots Ripton, Huntingdon, Cambridgeshire PE17 2LS, UK, e-mail p.rothery@ite.ac.uk; 3PO Box 2444, Mevasseret Zion 90805, Israel, frumkin@netvision.net.il
Newton, I., Rothery, P., & Frumkin, R. 1999. Post-fledging behaviour, dispersal and survival in Eurasian Sparrowhawks. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1397-1405. Johannesburg: BirdLife South Africa.Young male Sparrowhawks left the nest at about 26-27 days of age and young females at 28-30 days. The overall sex ratio among young at this age was equal, and did not vary with fledging date, brood-size and sex composition, age of mother, calendar year, or quality and elevation of nesting territory. For about four weeks after leaving the nest, the young remained in the nest vicinity, fed by their parents. On becoming independent, the young dispersed, males at an average age of 52 days and females at 55 days. At three nest-sites where food was experimentally provided beyond the end of the usual post-fledging period, young males stayed 16 days longer, and females 10 days longer, than young at control (unfed) nests. Some young from early nests, after dispersing from their natal area, joined younger broods still dependent on their parents, and thereby obtained additional food. From subsequent ring recoveries, young raised on high grade territories survived better through the post-fledging period than young on low grade territories. Survival also declined with increasing lateness of fledging date (within the season), but this trend was statistically significant only for females. Survival showed no significant relationships with brood size or sex composition, or elevation of nesting territory. Dispersal distances (judged from ringed birds reported dead) were greater in females than in males, and showed significant increases with increasing elevation of nesting territory and with lateness of fledging date. Young males from yearling mothers also dispersed further than young from older mothers. No significant relationships emerged between dispersal distances and grade of natal territory, brood-size or sex composition.
INTRODUCTION
In this paper, we describe briefly the behaviour of young Sparrowhawks Accipiter nisus in the post-fledging dependency period, and the influence of food-supply on the duration of that period. With the help of ring recoveries, we also assess the likely influence of the immediate post-fledging environment on post-fledging survival and dispersal.
Behaviour is analysed mainly from findings from 44 young that were radio-tagged, and followed through the post-fledging period until after they had dispersed away from the natal territory, while survival and dispersal were examined from the recoveries of nestlings ringed over a 27-year period. The radio-tagging work was conducted mainly in Rockingham Forest (Northants) in east-central England, while the ringing results came from two areas about 400 km to the north, Annandale and Eskdale in southern Scotland. In all three areas, Sparrowhawks were resident year-round; they nested in woodland, hunted in both wooded and open country, and ate almost entirely other birds. The work reported here formed part of a long-term study in all three areas (for further details, see Newton 1986, Wyllie & Newton 1991).
Young Sparrowhawks usually leave the nest for the first time at about four weeks of age (males a few days before females, see later) and for the next four weeks remain in the nest vicinity where they are fed by their parents. Throughout this period, the parents deliver food items to the nest, but as the young become more proficient flyers they increasingly take food on the wing directly from their parents. Towards the end of the post-fledging period, the parents often do not surrender the food immediately, but allow their young to chase them among the trees before releasing the prey. Until they disperse from the nest area, the young may continue to take some food items to the nest, which acts as a stable platform for feeding. On reaching independence, the young disperse abruptly and individually from the nest area, and in any direction.
A striking feature of the Sparrowhawk, like some other raptors, is the marked size difference between the sexes, with males smaller than females. This difference becomes increasingly pronounced during the course of the nestling period so that, by the age at which they are ringed, young Sparrowhawks can be sexed without error. This in turn enables the development, survival and dispersal of the two sexes to be examined separately. In adult life, males weigh only about half as much as females.
METHODS
During 1982-87, a total of 44 nestlings in 16 broods were individually marked with back-mounted radio-transmitters fixed by a diagonal loop harness, as described by Kenward (1985, 1987). Including the lithium/copper battery and the 25cm steel wire aerial, each transmitter weighed about 8 g, which was about 5% of the weight of a male Sparrowhawk and 3% of the weight of a female. Signals were detected with an AVM receiver (model LA12), using a hand-held three-element Yagi antenna (Kenward 1987). Throughout the post-fledging dependency period, each brood was checked every day, on a visit lasting 30-60 minutes, to record the numbers and positions of each young, and to listen for calls.If no bird was heard in an hour, a second visit was made on the next day before concluding that all birds had left that nest area. Whenever a radio-marked juvenile dispersed from the natal nest area, an attempt was made to follow it for several days (maximum 25), as it moved through the countryside. The analyses presented here incorporate the published records of Wyllie (1985) and Frumkin (1994), in which further details may be found.
To see whether food-supply had any influence on dispersal dates, additional food was provided to three broods of radio-tagged young in 1985.The food (mainly carcasses of Starlings Sturnus vulgaris and Quails Coturnix coturnix) was put on the nest at two sites and on a plucking post at the third. The sites were visited daily, the amount of remaining food was recorded, and additional food was provided. The amount of food supplied was always greater than the amount consumed the previous day, so the young should not have been hungry. Food was provided consistently from the time the young were 50 days old until they were about 80 days, some 20-30 days after the usual age of dispersal.
Over the years 1971-97, some 2845 young Sparrowhawks were ringed at nests in south Scotland. None of these were subsequently reported from the natal nest areas themselves in the post-fledging period. After their dispersal from their natal nest areas, some 215 ringed young were subsequently found dead and reported by members of the public, while 138 others were trapped alive as breeders in the study areas. To be recovered in either manner, both groups had to have survived through the post-fledging dependency period, so in order to assess survival through this period, both types of recoveries (live and dead) were pooled (total = 353), regardless of age at recovery (up to ten years). As a measure of dispersal, birds found breeding in the study areas gave only the shorter dispersal distances because they excluded any birds which bred outside the areas. In contrast, birds reported dead by members of the public should have given an unbiased picture of the directions and distances travelled. Hence, for the analyses in this paper, dispersal distances were taken only from birds found dead. No significant variations in recorded dispersal distances occurred between the different months of life from August on, at least to the end of the first year. This was in line with earlier conclusions, that the main dispersal period was short, and followed immediately after the post-fledging period. For our analyses of dispersal, therefore, recoveries were pooled from all months of the year.
The two response variables, namely survival through the post-fledging period and dispersal distances, were each examined in relation to:
(1) Elevation of nest above sea level (5-350 m).
(2) Grade (= quality) of natal territory (1 to 5, poor to good), based on previous occupancy and nest success (Newton 1991).
(3) Fledging date of the first young to leave the nest, estimated from laying date or from the growth stage of the young. In the population as a whole, dates of nest leaving were spread over about 45 days (nine successive 5-day periods).
(4) Brood size, from 1-6 at ringing (a few days before nest leaving).
(5) Sex composition of the brood as measured from (a) the proportion of males in the brood, and (b) whether the brood contained young of one sex or both sexes.
(6) Age of mother, whether in her first or subsequent years of life (recognisable from plumage, Newton 1986).
Data were initially analysed separately for each Scottish area, but are presented here only for the two areas combined, although allowing for any differences between areas.
RESULTS
The post-fledging period: behaviour, duration and dispersal
In the Sparrowhawk, the transition from nestling to post-fledging existence is gradual. When the young are about ready to fly, they walk onto branches near the nest or make short flights to nearby trees, scrambling back to the nest whenever a parent arrives with food. The young continue to feed on the nest throughout the post-fledging period, but in the latter half, they also take food on the wing directly from their parents, as mentioned above.
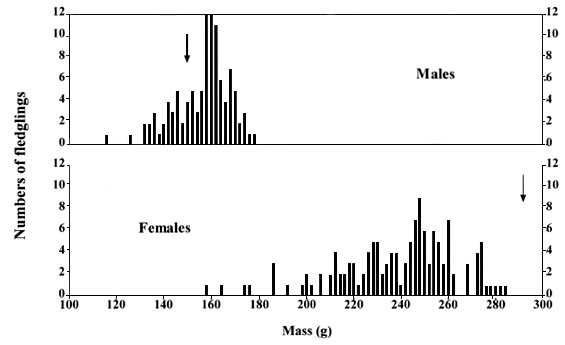
Young males left the nest at about 26-27 days of age and young females at about 28-30 days. On day 19-21, young males weighed an average of 157g ± SD 12g (n = 107), and young females about 53% heavier, at an average of 240g ± SD 25g (n = 133). In both sexes, individual variation in weight was great (Fig. 1), but whereas most young males were heavier than adult males at that time of year (mean 149 ± SE 2g)), young females were lighter than adult females (mean 289 ± SE 2g). Inspection of dead birds revealed that young males carried greater fat reserves than young females but no measurements were made.
During 27 years in the Scottish study areas, some 2845 nestlings in 803 broods were ringed, 1400 in 414 broods in Annandale (1971-81) and 1445 in 389 broods in Eskdale. Combining data from both study areas gave 1446 males and 1399 females, with a sex ratio, measured as the proportion of males, equal to 0.508 (SE = 0.009). No significant variation in sex ratio was found with brood size, laying date, year or any other of the variables examined.
Forty-four nestlings in 16 broods, which included 28 males and 16 females, were marked with radios, and checked daily thereafter, in order to ascertain the age of dispersal and subsequent movements. Within a few days, one of these birds (a female) was found dead, and three others (two males and a female) were not subsequently detected, which may have been because of radio failure or of predation and removal from the site. No other deaths or radio failures occurred until after dispersal, so mortality among the initial 44 young during the whole fledgling dependency period was 2.2-9.1%, depending on how many undetected birds had died. These losses left 40 birds (26 males and 14 females) which survived beyond the age of independence, and from which full data were obtained. During the post-fledging period, while still fed by their parents, the young gradually ranged over a wider area around the nest, until finally leaving the nest area, each on its own.
Thirty-one birds left abruptly in a single movement, and were not known to return during the study period, but the other nine (including three from experimental nests, see below) were recorded as far as 1.4 - 6.1 km (mean 3.4 ± SD 1.5 km) from the nest and then returned. Most of these young came back to the natal nest area within 24 hours of their first ‘dispersal’, but others spent 2-6 days away.Two categories of dispersal were therefore recognised: preliminary and ultimate (Frumkin 1994).
The mean age of preliminary dispersal was calculated at 52 and 55 days for males and females respectively, and of ultimate dispersal at 53 and 57 days. By this age, both sexes among these 40 birds had been out of the nest for an average of 27-28 days.
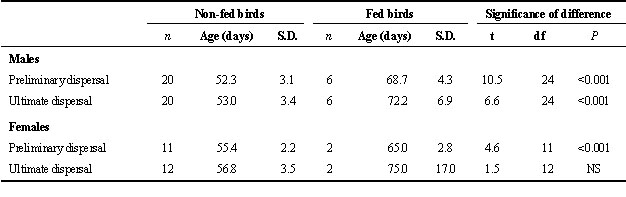
During the whole 4-week post-fledging period, young were not seen to catch food for themselves (though this may have happened unobserved). Because they gave more hunger calls (the ‘slow call’ of Frumkin 1994) towards the end of the post-fledging period, dispersal may have been triggered by reductions in the rate of prey delivery by their parents. It was to test this possibility that food was provided artificially to three nests for four weeks in 1985, beginning about one week before the young would have been expected to disperse. At these sites, preliminary dispersal occurred when the young were, on average 16.4 and 9.6 days older than non-fed males and females, respectively (Table 1). By the time food provision ceased, when the young were 80 days old, a few of the fed young had already left, while others stayed near the nest or returned to it for another 1-4 days, before finally leaving. Hence, for some of the young the dependency period, already prolonged by food provision, was cut short by our stopping food provision.The feeding experiment explained most of the variation in age of dispersal (for preliminary dispersal: r2 = 0.76, F1,38= 122, P<0.001; for ultimate dispersal r2 = 0.69, F1,38 = 82, P<0.001, both sexes combined), implying that food provision (by parents or observers) had a major influence on dispersal age. During many hours of observation, no aggression was seen either from parents to young or between young. Another obvious feature of the experimental young was that they almost never uttered food-begging calls. Delay in the age of dispersal was also observed in a similar food-provision study in Goshawks Accipiter gentilis (Kenward et al. 1993a).
On more than ten occasions in several different years, dispersing juvenile Sparrowhawks were caught at other nest-sites where broods were still present. Their ring numbers identified them as interlopers. Among the 40 radio-tagged birds, six from five early nests were recorded at 12 other nest-sites where young were still being fed by their parents. The distance of these sites from the natal nest ranged from 400m (which was the next nearest nest) to 6 km, and the time the birds were recorded on each site ranged from a few minutes to six successive days. On several occasions, such interlopers were seen chasing adults at those sites, while begging for food, and on other occasions they were seen feeding on the nest itself. These observations, together with the length of time that some interlopers spent with fledglings from another brood, left no doubt that they obtained food in this way, thereby extending their dependency period.
Post-fledging survival
Using ring recoveries, trends in post-fledging survival were measured as the proportions of birds in different categories that were found alive or dead after the end of the post-fledging period.
On the basis of ring recoveries, we used logistic regression to test for trends in survival with altitude, grade of territory, laying (reflecting fledging) date, brood size, sex ratio and age of mother (yearling verses older) (Table 2). We analysed trends for males and females separately in each area, and then again after combining records for both areas and sexes. In all analyses, allowance was made for any annual differences in recovery rates (expected because some annual cohorts had more years in which to be recovered than others), and in all analyses based on pooled data, allowance was also made for any differences in recovery rates between sexes and areas.
The most striking relationship was with grade of natal territory, in that birds of both sexes from good territories were more likely to be recovered after achieving independence than were birds from poor territories (Table 2). Other consistent trends were for declines in recovery rate with lateness of fledging date (significant only in males from both areas combined), with increasing elevation of nestling territory (not significant), and with lower age of mother (yearlings versus older birds, not significant). No other variables yielded consistent trends, and in particular, there was no evidence that brood size affected post-fledging survival. In an earlier analysis we could find no relationship within sexes between weight at fledging and subsequent survival (Newton & Moss 1986).
Post-fledging dispersal
In general, males dispersed less far (median distance 15.8 km) than females (median distance 29.5 km). Otherwise, most striking relationships were for dispersal distances to increase with lateness of fledging date (significant in males and both sexes combined using records for both areas), elevation of nesting territory (significant in females and both sexes combined, using records for both areas) and lower age of mother (yearlings versus older birds, significant for males and both sexes combined). No other significant relationships emerged (Table 3).
DISCUSSION
Despite the marked size difference between the sexes, male Sparrowhawks survived as well as females in the nest, causing no distortion in the hatching sex ratio by the time of fledging (Newton & Marquiss 1979). One reason for equal survival may be that the smaller males developed more rapidly, both behaviourally and physically, keeping ahead of their sisters throughout. The relatively greater weights and fat contents of males at fledging may have helped them to get through the post-fledging dependency period when the females caught up with them. The fact that, after independence, smaller proportions of males than females were recovered did not necessarily indicate differential survival: males were less likely to be found dead because they spent more time in wooded habitats than females (Marquiss & Newton 1982), and were less often caught alive at nests because they spent less time at nests than females (Newton 1986). Hence, nothing could be learnt about sex-related survival simply from the proportions recovered
The timing of dispersal, which marked the end of the post-fledging dependency period, did not appear to be influenced by parental aggression towards the young, nor by aggression among the young themselves (as none was seen), nor by a sudden cessation of food deliveries. However, it could have depended on a change in balance between parental provision and self provision. Towards the end of the dependency period, the parents gradually reduced their rate of prey deliveries (as noted in unpublished observations on radio-marked adults), and probably stopped once the flying skills of the young were adequate for them to hunt effectively. Yet in the food-provision experiment, the young stayed near the nests for longer than usual if their total food needs continued to be provided. Coincident with a reduction in the amount of food provided, the young ranged progressively further from the nest, and while away from the nest young might have occasionally managed to kill for themselves. Soon, their developing hunting skills may have allowed the young to stay away from the nest for longer periods, rather than wait near the nest for the parents to bring prey. At this stage, the parents themselves could stop feeding the young, and stay away from the nest. On this view, the timing of dispersal would have been influenced by an interaction between decreasing parental provision and increasing self provision. In the Spotted Flycatcher Muscicapa striata, Davies (1976) found that each fledgling became independent when the energy gain per unit effort of self-feeding exceeded the gain from begging and waiting for the parents.
The importance of this balance in the Sparrowhawk is shown by the fact that dispersing young would join other younger broods that were still being fed at a high rate. As Sparrowhawk parents could not discriminate between their own young and other young of similar age added to the nest at any stage of the nestling period (unpublished observations), it is unlikely that they could discriminate in the post-fledging period. One would not expect selection pressure to discriminate in the nestling period (because broods do not normally mix then); nor would one expect it in the post-fledging period because the parents dump most of the prey on the nest and can have no control over who eats it. They spend most of their time away hunting, and return to the nest area only to deliver food. Even when food is passed directly to a youngster, that youngster may often take it to the nest, feeding and leaving the rest for other young (usually its siblings). Moreover, even if parents do recognise intruders, the risks of killing offspring by mistake may inhibit attacks.
Such post-fledging brood-parasitism could clearly benefit the ‘parasitic’ young involved, helping them through the transition to full independence. It may be of disadvantage to the younger host brood, however, if some of the food that they might otherwise get is removed by the interlopers. The behaviour provides a clear selective advantage to the parents that breed early in the season, as their young are more likely to benefit from such behaviour than young raised later, while later young are more likely to act as hosts. Similar ‘nest switching’ behaviour by young fledged early in the season, has been recorded in the closely-related Goshawk Accipiter gentilis (Kenward et al. 1993b), and also in some other raptors, including Osprey Pandion haliaetus (Poole 1982), American Kestrel Falco sparverius (Lett & Bird 1987), Egyptian Vulture Neophron percnopterus (Donazar & Ceballos 1990), Red Kite Milvus milvus and Black Kite M. migrans (Bustamente & Hiraldo 1990).
Only 2-9% of 44 radio-marked young Sparrowhawks died during the post-fledging period itself, but all these young were on high grade territories. Post-independence recoveries from a much larger sample of Sparrowhawks ringed as nestlings indicated that the post-fledging environment influenced both survival and dispersal. For survival, the strongest relationship was with grade of territory. Earlier analyses revealed that all aspects of breeding were related to territory grade, with earlier laying dates, larger clutch and brood sizes and greater nest success on the higher grade territories (Newton 1991). Hence, post-fledging survival was simply another aspect of breeding that was better on higher grade territories. Post-fledging survival also declined with increase in lateness of fledging date, but the trend was significant only in males. This trend has been found in other species, notably Great Tit Parus major (Verboven & Visser 1998). Surprisingly, no relationship was found in Sparrowhawks between brood size and post-fledging survival, contrary to findings on some other species, such as Great Tit (Perrins 1979).
Another curious feature of Sparrowhawks was the lack of a relationship between fledgling body mass and subsequent survival, as discussed in an earlier paper (Newton & Moss 1986). This was also in contrast to findings on the Great Tit (Tinbergen & Boerlijst 1990, Verboven & Visser 1998).
For dispersal, the strongest relationship was with fledging date, with young raised late in the season moving further than those raised early. This could be interpreted in terms of dominance relations, with earlier (= older) young having priority of access to habitat nearest their natal areas, leaving later young to move further afield. This finding is paralleled in the Great Tit (Dhondt & Hublé 1968). In addition, young Sparrowhawks from yearling mothers tended to move further than those from older mothers and young from high elevation nests (= a poorer environment) tended to move further than young from low elevation nests.
Taken together, these findings imply that the post-fledging environment has considerable influence on the survival and dispersal of young Sparrowhawks. On the data available, we cannot say how much of the trends in survival occur as a result of differential mortality before independence, as opposed to after independence.
ACKNOWLEDGEMENTS
We are grateful to all landowners who gave us access to their woodlands, to Dr Mick Marquiss who contributed to the data collection in south Scotland, and to Professor C M Perrins for comments on the manuscript
.REFERENCES
Bustamante, J. & Hiraldo, F. 1990. Adoptions of fledglings by Black and Red Kites. Animal Behaviour 39: 804-806.
Davies, N.B. 1976. Parental care and transition to independent feeding in the young Spotted Flycatcher (Muscicapa striata). Behaviour 59: 280-295.
Donazar, J. & Ceballos, O. 1990. Acquisition of food by fledgling Egyptian Vultures Neophron percnopterus by nest-switching and acceptance by foster parents. Ibis 132: 603-607.
Dhondt, A.A. & Hublé, J. 1968. Age and territory in the Great Tit (Parus major L.). Angewandte Ornithologie 3: 20-24.
Frumkin, R. 1994. Intraspecific brood-parasitism and dispersal in fledgling Sparrowhawks Accipiter nisus. Ibis 136: 426-433.
Kenward, R.E. 1985. Raptor radio-tracking and telemetry. In Newton I. & Chancellor R.D. (eds) Conservation studies on raptors. I.C.B.P. Technical Publication 5: 409-420.
Kenward, R.E. 1987. Wildlife radio-tagging: field techniques and data analysis. London; Academic Press.
Kenward, R.E., Marcström, V. & Karlbom, M. 1993a. Post-nestling behaviour in Goshawks, Accipiter gentilis: I. The causes of dispersal. Animal Behaviour 46: 365-370.
Kenward, R.E., Marcström, V. & Karlbom, M. 1993b. Post-nestling behaviour in Goshawks, Accipiter gentilis: II. Sex differences in sociality and nest-switching. Animal Behaviour 46: 371-378.
Lett, D.W. & Bird, D.M. 1987. Post-fledging behaviour of American kestrels in southwestern Quebec. Wilson Bulletin 99: 77-82.
Marquiss, M. & Newton, I. 1982. Habitat preference in male and female Sparrowhawks. Ibis 124: 324-328.
Newton, I. 1986. The Sparrowhawk. Calton; Poyser.
Newton, I. 1991. Habitat variation and population regulation in Sparrowhawks. Ibis 133, suppl. 1.: 76-88.
Newton, I. & Marquiss, M. 1979. Sex ratio among nestlings of the European Sparrowhawk. American Naturalist 113: 309-315.
Newton, I. & Moss, D. 1986. Post-fledging survival of Sparrowhawks Accipiter nisus in relation to mass, brood-size and brood composition at fledging. Ibis 128: 73-80.
Perrins, C.M. 1979. British Tits. London; Collins.
Poole, A. 1982. Breeding Ospreys feed fledglings that are not their own. Auk 99: 781-785.
Tinbergen, J.M. & Boerlijst, M.C. 1990. Nestling weight and survival in individual Great Tits Parus major. Journal of Animal Ecology 59: 1113-1127.
Verboven, N. & Visser, M.E. 1998. Seasonal variation in local recruitment of Great Tits: the importance of being early. Oikos 81: 511-524.
Wyllie, I. 1985. Post-fledging period and dispersal of young Sparrowhawks Accipiter nisus. Bird Study 32: 196-198.
Wyllie, I. & Newton, I. 1991. Demography of an increasing population of Sparrowhawks. Journal of Animal Ecology 60: 749-766.
Table 1. Comparison of age at dispersal between control (non-fed) and experimentally fed young Sparrowhawks.
Table 2. Summary of tests for trends in survival with different variables related to the post-fledging period. The figures show deviance chi-square statistics for a trend using linear logistic regression, with sign indicating direction of trend. † P<0.10, *P<0.05, **P<0.01. Based on the pooled data for Annandale (1971-81) and Eskdale (1971-97) in south Scotland, but allowing for differences between years and between areas, and in the pooled data for males and females allowing also for differences between sexes.
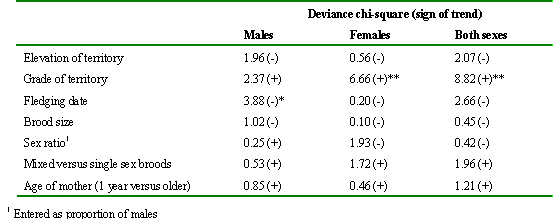
Table 3. Summary of tests for trends in dispersal distances (log distance) in relation to different variables. Figures show the percentage of variance accounted for by a linear trend after allowing for variation between areas and years, and in the pooled data for males and females allowing also for differences between sexes. † P<0.10, *P<0.05, **P<0.01.
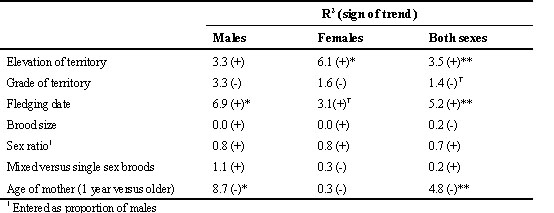
Fig. 1. Weights of young male and female Sparrowhawks at 24 days old, a few days before leaving the nest. Most males were heavier at this age than the average adult male (arrow) and females were lighter at this age than the average adult female (arrow). Data from Newton & Moss (1986).