S22.4: Conspecific attraction and breeding site selection in Kittiwakes: an experiment.
Thierry Boulinier1,2, Etienne Danchin2 & Stéphane Durand2
1 Department of Biology, Division of Zoology, University of Oslo, PO Box 1050, Blindern, N-0316 Oslo, Norway, e-mail tboulini@snv.jussieu.fr; 2 Laboratoire d’Ecologie, Université Pierre et Marie Curie, UMR 7625 CNRS, 7 Quai St Bernard, 75252 Paris, France, e-mail edanchin@snv.jussieu.fr
Boulinier, T., Danchin, E. & Durand, S. 1999. Conspecific attraction and breeding site selection in Kittiwakes: an experiment. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1315-1327. Johannesburg: BirdLife South Africa.Factors affecting the settlement of breeding individuals are likely to play an important role in the spatio-temporal distribution of breeding individuals. Thus, they are likely to influence the existence of aggregation of breeding territories, i.e. coloniality. The mere presence of conspecifics is known to affect settlement. Detailed observations in a Black-legged Kittiwake Rissa tridcatyla population moreover, suggested that the reproductive success of conspecifics (or correlates of RS) plays a prominent role in colony formation. Here we report the results of a small experiment carried out to test whether individuals would be more attracted to dummy nests containing dummy chicks than to empty dummy nests. Prospecting individuals were observed landing on these dummy nests and displaying ownership and sexual behaviours. The year after, several of the dummy nest-sites that contained dummy chick were occupied by pairs that raised nestling. The results suggest that the content of nest, as well as their location relative to other evidence of local successful breeding activities attract prospectors who may recruit in the following year. These findings have implications for both conservation, and the understanding of the evolution of coloniality.
INTRODUCTION
Among the various hypotheses proposed to explain the evolution of coloniality, several discuss the balance of costs and benefits associated with breeding in groups of different sizes, but no clear answer has emerged from this approach (Wittenberger and Hunt 1985, Shields et al. 1988, Danchin and Wagner 1997). Assessing the balance of costs and benefits of breeding within a given colony size is difficult because some factors given as benefits of breeding aggregation - such as better defence against predators, local availability of mates etc. - can also be regarded as costs - respectively, increased attraction of predators, risk of losing paternity etc. (Danchin and Wagner 1997). One way of solving the question of why some bird species form aggregations of breeding territories, may be to consider the habitat selection processes that may lead individuals to form such aggregations. Recently, the hypothesis that coloniality evolved as a by-product of habitat selection mechanism based on conspecific reproductive performance has been proposed following detailed studies carried out on Black-legged Kittiwakes Rissa tridactyla (Boulinier and Danchin 1997, Danchin et al. 1998). This hypothesis is based on the idea that places where there are conspecifics are likely to be suitable (Stamps 1988), and that places where conspecifics are performing well are even more likely to be good (Shields et al. 1988), thus leading to dynamic aggregations of breeding individuals among potential patches (Boulinier and Danchin 1997, Danchin et al. 1998). This mechanism of habitat selection can work if the potential breeding patches varies strongly in quality and if local patch quality is sufficiently predictable (Boulinier and Danchin 1997).
The Kittiwake is a relatively long-lived pelagic seabird which breeds on vertical cliffs. Nest building and chick rearing duties are shared by both members of the pair. Most individuals first breed when three or four years-old (Danchin and Monnat 1992), and younger individuals are usually less successful than older ones (Coulson and Thomas 1985). As in most long-lived birds, prospecting birds, defined as birds not breeding locally this current season, are observed in the colonies at the end of the breeding season (Porter 1988, Porter 1990, Danchin et al. 1991, Monnat et al. 1991, Cadiou et al. 1994). These prospectors appear to be attracted to the most productive parts of the breeding colonies (Danchin et al. 1987, Cadiou, in press) and the timing of prospecting corresponds to the timing of availability of reliable information on the relative quality of the different breeding patches within the colonies (Boulinier et al. 1996). A particular category of prospecting individual has been observed ‘squatting’ temporarily on nests left unattended by breeders (Danchin et al. 1987, Monnat et al. 1990, Cadiou et al. 1994). Unattended nests are notably squatted by the older prospecting birds, which are the most likely to recruit locally in the following year (Cadiou et al. 1994). Prospecting and the related ‘squatting behaviour’ have been observed in other cliff nesting (e.g., in the Black Noddy, Cullen and Ashmole 1963) and hole nesting seabirds (e.g., in the Atlantic Puffin Fratercula arctica, Ashcroft 1972), and are suspected of playing an important role in the process of local recruitment of individuals to highly productive breeding patches (Monnat et al. 1990, Danchin et al. 1991, Reed et al. in press). Such potential processes may thus have important consequences for the dynamics of sub-populations linked by dispersal events, as well as for the understanding of coloniality. It is nevertheless not clear whether birds actually cue on local reproductive success (or some social correlates) or directly on the local factors that may potentially affect predictably the quality of the local breeding patch (Boulinier and Lemel 1997). In this context, we planned an experiment in an attempt to test whether the presence of artificial nests and decoys of chicks on an unoccupied portion of cliff could attract prospecting individuals and ultimately lead to local recruitment. It was predicted that prospecting individuals would be seen more frequently on the section of the cliff with a higher proportion of artificial nests containing decoy chicks, and that they would display more behaviour related to pair formation and site acquisition on these parts of the cliff. It was also predicted that birds that should be observed displaying such behaviours on the artificial cliff would be either individuals that have not bred before (pre-breeders), or that have bred in the past but did not breed that given year (‘sabbatical’, Cadiou et al. 1994), or current breeders that failed their breeding attempt.
METHODS
The study site
The study was done in the context of an ongoing long-term study of the population dynamics and behaviour of Kittiwakes carried out at the Cap Sizun, on the western tip of Brittany, France (Danchin and Monnat 1992, Danchin et al. 1998). The breeding population at the Cap Sizun averaged approximately 900 pairs in the several years preceding the experiment, with much local variation occurring in the number of breeders (Danchin and Monnat 1992). The small size of the population, an extensive colour banding of individuals (especially of nestlings), and frequent observations since 1979 provided information on the history and current breeding status of some individuals potentially observed prospecting on the artificial cliff. In this population, the breeding season spans from January to August, with the incubation period in May and most of the fledging period occurring during the second half of July (Cadiou et al. 1994). From the end of June, a decrease in the attendance of breeders at the nest is usually observed, and at that period individual prospecting birds are seen squatting on nests with nestlings (Cadiou et al. 1994).
The experiment
In 1994, a small cliff (10 meters x 10 meters) adjacent to an existing colony situated outside the Cap Sizun Nature Reserve was chosen as the experimental site (Fig. 1). The cliff, chosen to represent a potentially suitable breeding patch for Kittiwakes, had only hosted one Kittiwake nest since the creation of the colony in 1983, and was not currently holding any breeding attempts. It was relatively less steep than most of the nearby occupied cliffs, and may not have represented an optimal breeding patch for Kittiwakes. On its south section (left section), it was adjacent to a small breeding cliff occupied by successfully breeding Kittiwakes, and on its north section (right section), it was adjacent to a ‘club’, where young pre-breeders and of-duty breeders commonly gather.
The ‘experimental cliff’ was divided in four portions (sub-patches), within which a group of 10-11 artificial nests was set at the beginning of July 1994 (Fig. 1). Two sub-patches contained mostly ‘successful’ nests, and the two other contained mostly ‘unsuccessful’ nests (Fig. 1). The location of the successful sub-patches was determined randomly, with the constraint that they should not both be situated in the upper or lower parts of the cliff. Artificial nests were constructed of dry grass and plaster. Nests were attached to the cliff with plaster and the natural local topography. We placed decoys of chicks in half of these nests. Kittiwake chicks have a plumage quite different from the one of the adults; we thus drew the chicks in black on 2D white polystyrene cardboard which had been cut to the shape of the profile of a chick displaying the appeasement posture. When displaying the appeasement posture, the black bar on the back of the neck of the nestling is displayed and the bill is hidden down on the chest. We did not expect the birds to actually take the dummy chicks for actual nestlings after landing on the artificial nest site, but we believe that such dummy chicks seen from a distance could be taken for actual chicks left unattended in their nest. The use of decoys and 2D images of conspecifics has been used successfully in both laboratory studies of recognition in domestic birds (e.g., D’Eath and Dawkins 1996), and in ecological studies of conspecific attraction in seabirds (e.g., Podolsky 1990).
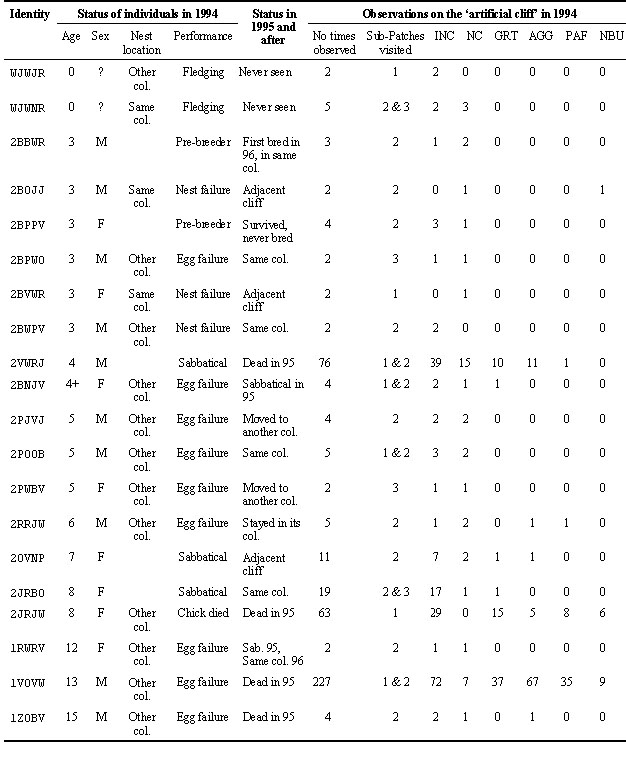
Prospecting is known to increase as the season progresses (Cadiou 1993, Cadiou et al. 1994, Boulinier et al. 1996), and the date for setting up the nests on the experimental cliff was thus chosen to correspond to the beginning of the active prospecting period. The attendance and behaviour of birds landing on sites of the cliff were then recorded for an average of three hours per day on 26 days. The identity of the birds was recorded for colour banded individuals. Behaviour was recorded according to a detailed ethogram of the Kittiwake (Danchin 1987a and b, 1991), and then grouped into several main functional categories. This permitted the estimation of the frequency of behaviour related to site acquisition and pair formation on the different sites and portions of the experimental cliff. The frequency of behaviour (number of times the behaviour was recorded per unit of time of observation) was computed per group of sites and per day. In order to test for a potential effect of the distance between the site and the nearby actual breeding colony, sites were ranked according to distance to the closest active nest containing nestlings.
RESULTS AND DISCUSSION
Which birds visited the cliff?
Several individuals visited the study plot over the prospecting period. Most of the landings occurred on sites with dummy nests (59% over a total of 1214). The right side of the study plot was adjacent to a ‘club’ where birds spend time resting, and most birds that were recorded on that part of the plot may thus have been only resting rather than prospecting on the cliff. This was notably the case for individuals that landed on sites with no dummy nest. Most birds recorded on other parts of the cliff nevertheless displayed behaviours associated with prospecting of nest sites (Table 1 and see below).
Visits of 20 colour marked individuals to the study plot were recorded over the study period. These observations, combined with what was known about the breeding history of the birds, permitted us to get an idea about the status of the birds that visited the cliff. None of the birds recorded on the cliff bred successfully in 1994 (Table 1). Two of them were young of the year that had just fledged, but all of the others were old enough to have bred successfully that year. Two of the individuals were from a pair that failed breeding on a site nearby the artificial cliff within the same colony, and all other individuals had either not bred that year (2 pre-breeders and 3 sabbatical individuals), or failed their breeding attempt on other colonies than the one that contained the artificial study cliff (Table 1). Despite the small size of the sample of marked birds, this shows that the artificial cliff had been, as expected, visited by failed breeders and non-breeders, i.e. birds potentially seeking a breeding site for a future season.
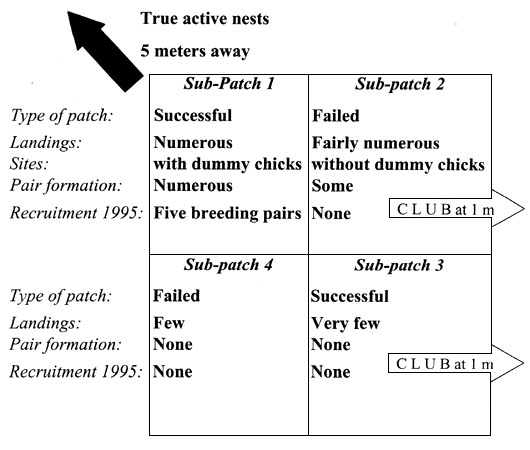
As will be seen below, active prospecting behaviour of individuals was mostly observed on the top-left sub-patch, where four dummy nests containing dummy chicks attracted several individuals. Recorded behaviours were not independent of each other, notably as some individuals showed temporary attachment (over several days) to some of the sites and got involved in several displays and interactions that were recorded. Over the 26 day period, it appeared nevertheless clear that several individuals had been involved, thus diminishing that problem of non-independence.
Attendance and behaviour of birds prospecting on the cliff
The distribution of landings and of behaviour of birds within the cliff showed strong heterogeneity. As mentioned above, the majority of landings observed occurred on dummy nests, apart from one of the 4 sub-patches where 323 of the 329 landings did not occur on dummy nests: most of these were due to birds actually resting on the border of the adjacent ‘club’, and could not be considered as prospectors attracted to the cliff. For this reason, the observations of individuals that landed outside of dummy nests were not included in the analyses on the relative attractiveness of different parts of the artificial cliff. The overall distributions of the different categories of behaviour observed on (1) sites with no dummy nest, (2) sites with a dummy nest only, and (3) sites with a dummy nest containing a dummy chick, show clearly that active prospecting occurred on the dummy nest sites, but not much off these sites (Table 2).
The mean number of landings recorded per unit of time differed among sub-patches within the artificial cliff (ANCOVA, main effect of the sub-patches: F 3, 61 = 16.80; P < 0.0001), and increased through time (main effect of the day of observation: F 1, 61 = 36.13, P < 0.0001; Fig. 2). We recorded a higher mean number of landings on dummy nests on the higher half part of the artificial cliff (sub-patches 1 and 2) than on the lower part (sub-patches 3 and 4; F 1, 61 = 31.02, P < 0.0001). The two lower sub-patches may have actually been too low (just few meters above sea level) to be considered by the birds as containing actual potential breeding sites. We did not record a higher mean number of landings on dummy nests on the left part of the artificial cliff than the right part (F 1, 61 = 2.83, P = 0.0974).
The frequency (occurrence per unit time) of the various categories of behaviour varied according to the sub-patch and time. Behaviour included in the category of pair formation (PAF) and site acquisition (e.g., greetings, GRT) were recorded at higher frequencies on the top left ‘successful’ sub-patch than on the other sub-patches (ANCOVAs with day of observation as a continuous variable; sub-patch effect for PAF: F 1,138 = 6.28, P = 0.0134; for GRT: F 2,219 = 9.99, P = 0.0001; numbers of degrees of freedom differs from 3 as some behaviour was not observed on some of the sub-patches). These two categories correspond to behaviour that is displayed on an actual site that may hold a potential future breeding attempt. It is thus noticeable that they were recorded more frequently on this sub-patch containing the dummy nests with dummy chicks. Aggressive behaviour (AGG) and confident but inactive behaviour (INC) were also recorded at higher frequencies on this top left sub-patch (For AGG: F 2,262 = 3.19, P = 0.0439; for INC: F 3,845 = 84.75, P = 0.0001). Conversely, the frequency of non-confident behaviour (NC) did not differ among sub-patches (F 3,301=2.27, P = 0.0805). Evidence of an association between the presence of dummy chicks in nests and the behaviour of the prospecting birds were apparent when we looked at the direct effect of the content of dummy nest (empty versus with a dummy chick; for PAF: F 1,139 = 6.28, P = 0.0134, for GRT: F 1,220 = 13.82, P = 0.0003). When the distance with the very nearby active breeding cliff was controlled for (as a continuous variable in the ANCOVA), there was no longer an effect of the presence or absence of dummy chicks in the nests: the presence of the nearby active group of breeding Kittiwakes potentially confound a possible causal association between the content of the dummy nest and the frequency of the prospecting behaviour recorded. It is nevertheless noticeable that it is the presence of actual successful breeders nearby that may have participated to the attractiveness of the group situated on the top-left sub-patch of the artificial cliff.
Local recruitment the year afterNone of the marked birds observed on the artificial cliff in 1994 recruited to that cliff in 1995. The fate of these birds in 1995 nevertheless shows that they were in the process of changing site at the time they were observed on the artificial cliff in 1994 (Table 1): among the 11 individuals that bred in 1995, 2 changed nest site within the colony where the artificial cliff was set up, and 5 locally recruited to that colony. Four of these birds actually dispersed from a colony situated about 20 kilometres away, where they had failed their reproduction in 1994 at the egg stage when heavy predation by Ravens caused massive local failures.
In 1995, five nests were built by birds on the experimental plot. Three were on former experimental nest sites with dummy chicks, two on sites very close to experimental sites with dummy chicks. Four more nests were built in 1996. In 1997 and 1998 again, nine sites were occupied. A total of 10 sites have been used on that cliff since the experiment, of which seven corresponded to experimental sites (all with Dummy chicks) and three to close ledges. Over the same period, the colony within which the artificial cliff was situated increased from 64 nests in 1993 to 435 in 1997. This large increase involved heavy immigration associated to differential local reproductive success among colonies, and must be considered when looking at the local recruitment on the ‘artificial’ cliff. However, it is noticeable that although that colony had hosted similar numbers of nests at the end of the 1980’s (see Fig. 3), only one nest had ever been recorded on the artificial cliff.
In summary, this small experiment showed that (1) potential prospectors could be effectively attracted to a cliff where artificial nests were set up and, (2) that the distribution of landings on that artificial cliff was not random (Fig. 5), and may have been affected by the nest content (empty versus containing a dummy chick), their distance to a nearby active cliff and their height.
The lack of an actual replicated design and the very artificial nature of the dummies used as local evidence of reproductive success nevertheless limit the inferences that can be made from the results of this experiment. For instance, the dummy chicks may have served as evidence of local successful reproduction when individuals first encountered the cliff from a distance, but may not have played a significant role thereafter. It is nevertheless interesting to note that the birds mostly displayed ownership and sexual behaviour on nests containing dummy chicks and that most nest sites that eventually got occupied the year after had had a dummy chick. These results match detailed behavioural observations made in the Kittiwake (Danchin 1987, Cadiou et al. 1994), notably the fact that individuals are most likely to recruit to one of the sites where they have been observed squatting on chicks. These results are also in accordance with the work of Kress and collaborators (Podolsky and Kress1989) that have been using different decoys and devices to attract potential breeders of different seabird species for conservation purposes: they notably observed that a plot with decoys of adult Common Guillemots Uria aalge with eggs attracted more individuals than a plot with adults with no eggs (Kress 1993). Seemingly, Slagsvold and Lifjeld (1990) found that return rates of yearling male Pied Flycatchers were higher in a study plot with higher local reproductive success than in a plot were local success was experimentally reduced. Due to the large spatial and temporal scales involved, the few experimental studies dealing with the potential effect of local reproductive success of conspecifics on recruitment and dispersal usually lack proper replicates. It is nevertheless clear that only an experimental approach can determine whether the local reproductive success of conspecifics (per se or some social correlates) directly affects dispersal and recruitment processes, and thus the functioning of fragmented bird populations (Switzer 1997). Our results suggest that much insight can be gained by following the behaviour of known individuals after a local manipulation of breeding success. They also underline the importance of considering the dynamics of recruitment and dispersal at different scales, in order to predict the local dynamic with breeding colonies. The understanding of these processes has strong implications in conservation biology, and also for the study of the evolution of coloniality (Boulinier and Danchin 1997, Danchin and Wagner 1997): colonies may need to be viewed more as dynamic and heterogeneous assemblages of breeding individuals than as relatively static groups of breeding birds.
ACKNOWLEDGEMENTS
We thank Valérie Audinot for her help in the field and Mathieu Guillemin for help with data management and analysis. Richard Wagner provided judicious comments and improvements to the manuscript.
REFERENCES
Aschroft, R.E
. 1976. Breeding biology and survival of Puffins.PhD Thesis, Oxford.Boulinier, T. & Lemel, J.-Y. 1996. Spatial and temporal variations of factors affecting breeding habitat quality in colonial birds: some consequences for dispersal and habitat selection. Acta Oecologica 17: 531-552.
Boulinier, T. & Danchin, E. 1997. The use of conspecific reproductive success for breeding patch selection in territorial migratory species. Evolutionary Ecology 11: 505-517.
Boulinier, T., Danchin, E., Monnat, J.-Y., Doutrelant, C. & Cadiou, B. 1996. Timing of prospecting and the value of information in a colonial breeding bird. Journal of Avian Biology 27: 252-256.
Cadiou, B. 1993. L’accession à la reproduction: un processus social d’ontogénèse. Cas de la mouette tridactyle (Rissa tridactyla). PhD Thesis, Université de Rennes I, France.
Cadiou, B. 1998. Attendance of breeders and prospectors on cliffs reflect the quality of breeding places in the Kittiwake Rissa tridactyla. Ibis, in press.
Cadiou, B. & Monnat, J.-Y. 1996. Parental attendance and squatting in the Kittiwake Rissa tridactyla during the rearing period. Bird Study 43: 164-171.
Cadiou, B., Monnat, J.-Y. & Danchin, E. 1994. Prospecting in the Kittiwake, Rissa tridactyla: different behavioral patterns and the role of squatting in recruitment. Animal Behaviour 47: 847-856.
Coulson, J.C. & Thomas, C. 1985. Differences in the breeding performance of individual Kittiwake gulls, Rissa tridactyla (L.). In: Sibly, R.M. & Smith, R.H. (eds). Behavioural Ecology. Oxford, Blackwell Scientific Publications: 489-503.
Cullen, J.M. & Ashmole, N.P. 1963. The Black Noddy Anous tenuirostris on Ascension island. Part 2. Behaviour. Ibis 103b: 423-446.
Danchin, E. 1987a. The behaviour associated with the occupation of breeding site in the Kittiwake gull Rissa tridactyla: the status of landing birds. Animal Behaviour 35: 81-93.
Danchin, E. 1987. Les comportements liés à l'occupation du site de reproduction chez la Mouette tridactyle Rissa tridactyla. Les comportements de pré-atterrissage. Behaviour 100: 226-246.
Danchin, E. 1991. Social displays in the Kittiwake Rissa tridactyla. Bird behaviour 9: 69-80.
Danchin, E. & Monnat, J.-Y. 1992. Population dynamics of two neighbouring Kittiwake Rissa tridactyla colonies. Ardea 80: 171-180.
Danchin, E. & Wagner, R. 1997. The evolution of coloniality: the emergence of new perspectives. Trends in Ecology and Evolution 12: 342-347.
Danchin, E., Monnat, J.-Y. & Pasquet, E. 1987. Rôle des facteurs comportementaux dans les mécanismes de régulation des populations d’oiseaux coloniaux. Actes du Colloque National du CNRS ‘Biologie des Populations’: 618-627.
Danchin, E., Boulinier, T. & Massot, M. 1998. Habitat selection based on conspecific reproductive success: implications for the evolution of coloniality. Ecology, in press.
Danchin, E., Cadiou, B., Monnat, J.-Y- & Rodriguez Estrella, R. 1991. Recruitment in long-lived birds: conceptual framework and behavioural mechanisms. Proceedings of the. International Congess of Ornithology 20: 1641-1656.
D’Eath, R.B. & Dawkins, M.S. 1996. Laying hens do not discriminate between video images of conspecifics. Animal Behaviour 52: 903-912.
Kress, S.W. 1993. More murres on Matinicus rock. Egg Rock Update - Unpublished report.
Monnat, J.-Y., Danchin, E. & Estrella, R. 1990. Assessment of environmental quality within the framework of prospection and recruitment: the squatterism in the Kittiwake. Comptes Rendus de l’Académie des Sciences de Paris (III): 391-396.
Podolsky, R.H. 1990. Effectiveness of social stimuli in attracting Laysan Albatross to new potential nest sites. Auk 107: 119-125.
Podolsky, R.H. & Kress, S.W. 1989. Factors affecting colony formation in Leach’s storm petrel. Auk 106: 332-336.
Porter, J.M. 1988. Prerequisites for recruitment of Kittiwakes Rissa tridactyla. Ibis 130: 204-215.
Porter, J.M. 1990. Patterns of recruitment to the breeding group in the Kittiwake Rissa tridactyla. Animal Behaviour 40: 350-360.
Reed, J.M., Boulinier. T., Danchin, E. & Oring, L. In Press. Informed dispersal: prospecting by birds for breeding sites. Current Ornithology, Volume 15.
Shields, W.M., Crook, J.R., Hebblethwaite, M.L. & Wiles-Ehmann, S.S. 1988. Ideal free coloniality in the swallows. In Slobochikoff, C.N. (ed). The ecology of social behaviour. San Diego, Academic Press: 189-228.
Slagsvold, T. & Lifjeld, J.T. 1990. Return rates of males pied flycatchers: an experimental study manipulating breeding success. In: Blondel, J. (ed.). Population biology of passerine birds. NATO ASI Series 24, Berlin, . Springer-Verlag: 441-452.
Stamps, J.A. 1988. Conspecific attraction and aggregation in territorial species. American Naturalist 131: 329-347.
Switzer, P. 1997. Past reproductive success affects future habitat selection. Behavioral Ecology and Sociobiology 40: 307-312.
Table 1. Breeding status and summary of behavioural observations of the marked birds that visited the artificial cliff. Categories of behaviour reported are: INC: Inactive, confident; NC: Non-confident; GRT: Greetings and other ownership displays; AGG: Aggressive displays; PAF: Pair formation activities; NBU: Nest building activities. Same colony: colony containing the experimental cliff; Adjacent cliff: other cliffs than experimental cliff within experimental colony; Sabbatical: non-breeder
Table 2. Distribution of the total number of different categories of behaviour observed on three types of sites. Categories of behaviour are as in legend of table 1.
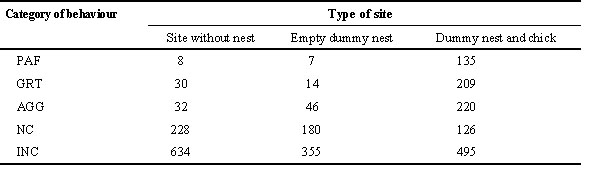
Fig. 1. Map of the cliff where artificial nests and decoys of chicks were set. Black dots for dummy nests with dummy chicks; open dots for dummy nests without dummy chicks; X for close sites that were occupied at least once by visiting birds. True active nests were at a few meters from the top left corner.
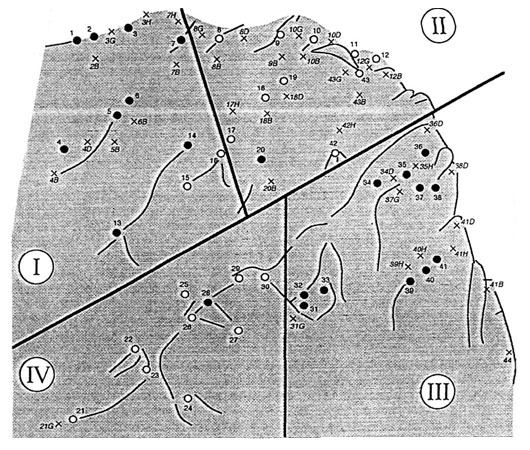
Fig. 2. Frequency of landings on dummy nests of the four sub-patches through time.
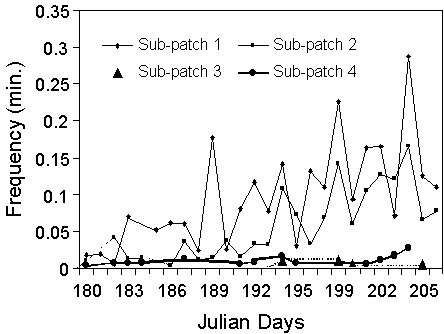
Fig. 3. Numbers of breeding pairs in the total colony and in the artificial cliff since the onset of breeding in that colony in 1983 (the arrow indicates the year the artificial nests and decoys of chicks were put on the artificial cliff)
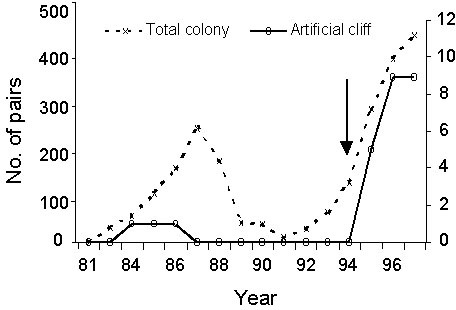
Fig. 4. Frequency of ownership and sexual behaviour in the four sub-patches through time.
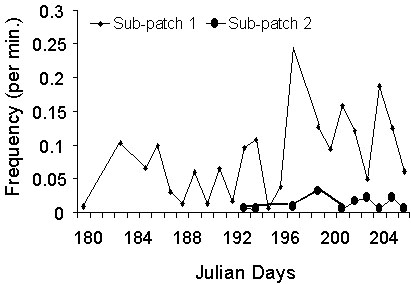
Fig. 5. Summary of the results obtained on the four sub-patches. The diagram respects the relative spatial position of the sub-patches.