S22.1: The Evolution of coloniality: The importance of colony size
Charles R. Brown & Mary B. Brown
Department of Biological Sciences, University of Tulsa, 600 S. College Ave., Tulsa, OK 74104-3189, USA, e-mail: charles-brown@utulsa.edu
Brown, C.R. & Brown, M.B. 1999. The evolution of coloniality: The importance of colony size. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1293-1303. Johannesburg: BirdLife South Africa.The size of breeding colonies varies widely in most colonial birds. This natural size variation has allowed an understanding of how the costs and benefits of coloniality change with group size, but the reasons for variation in colony size are unknown. We suggest that colony-size variation offers insights into the evolution of coloniality. If colony size is proportional to local resource availability (an ideal free distribution), each individual regardless of its breeding group should receive an equal fraction of the resource and have similar fitness expectations. In such cases, reproductive success should not vary with colony size. Field data for herons suggest that colony size directly reflects local resource abundance, but little comparable data for other species exist. If colony size reflects an ideal free distribution, coloniality likely evolved in response to resource heterogeneity within a habitat. Colony size may also reflect phenotypic differences among birds (a modified ideal free distribution), with birds of particular ages or abilities sorting into colonies of different sizes based on the costs and benefits of group size that each individual expects. If composition of colonies differs, different birds may be receiving different benefits from colonial breeding with colony size reflecting individual optima. Arguments that colony size is unstable and thus cannot be a primary determinant of fitness assume a particular relationship between fitness and group size, that an optimal colony size exists, and that choice of colony size is done individually (not through collective processes). None of these assumptions is strongly supported by data, and some are violated for some species, implying that colony size may be stable at times. Field workers should concentrate on determining why colonies vary in size.
INTRODUCTION
Most colonially breeding birds show extensive variation in colony size within populations. This variation can be over several orders of magnitude and is seen in virtually all taxonomic groups (Brown et al. 1990). Field studies of avian coloniality have used natural colony-size variation to explore many of the classical costs and benefits of group living (e.g., Hoogland and Sherman 1976; Veen 1977; Robinson 1985; Brown and Brown 1986, 1987, 1996; Van Vessem and Draulans 1986; Møller 1987; Shields and Crook 1987; Burger and Gochfeld 1990, 1991; Wiklund and Andersson 1994), and these studies have revealed that costs such as ectoparasitism and benefits such as avoidance of predators tend to vary systematically with colony size. In the best studied case, that of the Cliff Swallow Petrochelidon pyrrhonota at least 13 different costs and 13 different benefits were identified, with the magnitudes of these dependent to some degree on colony size (Brown and Brown 1996). While a cost-benefit analysis of these factors is complicated by their being measured in different currencies (for example, the probability of adult survival versus the incidence of food piracy by kleptoparasitic fledglings), clearly colony size can influence an individual's fitness. Unclear, however, is why colonies vary in size. Although a few hypotheses have been offered (Brown et al. 1990), virtually no field studies have addressed the ecological or social bases for variation in breeding colony size.
This paper is not a review of the extensive literature on the costs and benefits of coloniality; such reviews are available from Burger (1981), Birkhead (1985), Wittenberger and Hunt (1985), Brown et al. (1990), Siegel-Causey and Kharitonov (1990), and Brown and Brown (1996). Instead, we focus on what colony size may tell us about the evolution of avian coloniality. We have argued elsewhere (Brown and Brown 1996) that an explanation of colony-size variation will likely explain the evolution of coloniality more generally, and we expand on that argument here.
IDEAL FREE DISTRIBUTIONS AND LOCAL RESOURCES
That colony size might reflect an ideal free distribution of birds with respect to patchy resources within a habitat has been essentially the only widely discussed hypothesis for colony-size variation (Shields et al. 1988; Brown et al. 1990; Brown and Rannala 1995; Brown and Brown 1996; Danchin and Wagner 1997). This view holds that local variation in resources or intrinsic habitat quality lead to occupancy by birds in direct relation to resource abundance. This is the general basis for Danchin and Wagner's (1997) habitat-selection model for colony formation. Resources may be commodities such as amount of food or number of nesting sites and potential mates, or aspects of nesting habitat quality such as relative degree of predation or ectoparasitism experienced. The net effect is that more birds settle in areas of high abundance and fewer birds in areas with low resource abundance. If the distribution is truly ideal free (with continuous input of resources), each individual regardless of where it settles receives an equal share of the total resources in the habitat as a whole. Colony-size variation would then reflect variation in intrinsic quality of habitat patches.
The pervasiveness of colony size variation among species (Brown et al. 1990) might suggest that birds commonly distribute themselves among patches in an ideal free manner. Although ideal free theory has been well studied (e.g., Fretwell and Lucas 1970; Sutherland and Parker 1985, 1992; Parker and Sutherland 1986; Houston and McNamara 1988) and the concept applied frequently to foraging groups of different sizes (Milinski 1988; Tregenza 1995), no one has tested directly whether breeding colonies represent ideal free distributions with respect to habitat quality. Brown and Rannala (1995) indirectly tested whether Cliff Swallow colonies reflect an ideal free distribution by comparing observed colony sizes at sites across years to a theoretical distribution based on a Poisson process. Making the assumption that resources at a site are autocorrelated between years, they found that swallows were not distributed among sites in an ideal free way. This approach has promise and should be applied to other species, but it is informative only when empirical results differ from the theoretical distribution (rejecting the ideal free distribution). If data fit the model, the ideal free distribution cannot be confirmed because other processes may generate the same distribution of colony sizes.
We suggest two alternative ways to address whether colony sizes reflect ideal free distributions in colonial birds. One is to measure local resources directly. This requires identifying the relevant resource(s) in a habitat patch, which itself is a major challenge because so many potentially affect colony size (Brown and Rannala 1995; Danchin and Wagner 1997). More total resources should be associated with larger colonies. Perhaps the most obvious resource to influence colony size in this way is food. There is, however, relatively little information available on food abundance at colonies of different sizes. Part of this reflects the difficulty of measuring resources on the spatial scale required; colonial seabirds, for example, feed over large areas, and sampling food on such a scale is prohibitively difficult (Hunt and Schneider 1987, Cairns 1992). In species such as Cliff Swallows which feed on many different kinds of prey, sampling them all is virtually impossible because of the diversity of methods required (Brown and Brown 1996).
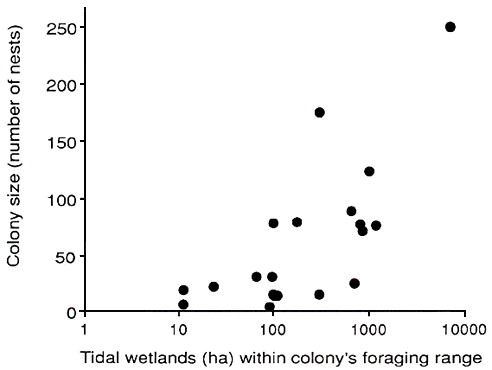
An alternative approach for some species is to assess the amount of foraging habitat near colonies of different sizes, assuming that a greater foraging area supports more food. This method was used in studies of Great Blue Herons Ardea herodias in Maine and Illinois (Gibbs et al. 1987; Gibbs and Kinkel 1997). In both populations, amount of preferred foraging habitat within a colony's foraging range correlated directly with colony size at a site (Fig. 1). Heron colony size directly reflects the amount of locally available food, and Gibbs' studies provide the best demonstration yet of colony size as an ideal free distribution of birds with respect to resources. This result, if general, would imply that colony formation occurs primarily in response to patchy and uneven resource distributions. While the possible influence of patchy resources on the evolution of coloniality has been recognized for decades (e.g., Crook 1965; Horn 1968; Alexander 1974) and assumed in recent discussions (Danchin and Wagner 1997), to date the Great Blue Heron data are the only convincing ones that show a direct link between extrinsic features of a habitat patch and colony size.
The other way to study the ideal free distribution is to use reproductive success. If birds distribute themselves such that each receives an equal fraction of the total resource, each individual should have a similar breeding success. We would predict under these conditions, therefore, that average fitness should not vary with colony size. A recent review (Brown and Brown 1996) found 16 studies that showed no relationship between annual reproductive success and colony size, supporting an ideal free process in the evolution of coloniality. However, the same review found over 30 other studies which showed a positive, negative, or curvilinear relationship between annual reproductive success and colony size. The evidence from studies of reproductive success alone thus does not support the hypothesis that avian colony size in general reflects ideal free use of resources. We emphasise, though, that only the approach of Gibbs et al. (1987) and Gibbs and Kinkel (1997) or other direct measures of resource availability can address the issue conclusively.
In assessing any relationship between colony size, local resources, and expected fitness, it is important to distinguish to what degree habitat quality causes colony size and to what degree it reflects the consequences of colony size. The choice of a breeding site occurs either during the previous nesting season often during chick rearing (Boulinier et al. 1996) or early in the current breeding season (Brown and Brown 1996). Resource availability in a habitat patch at these times would be the critical determinant of how many individuals can settle and what size colony forms there. Initial settlement could be based on an ideal free process, and if resources were measured at that time, one should find colony sizes proportional to resource availability. However, not all of the social costs and benefits of group living are linearly related to colony size; some operate more strongly above or below certain colony-size thresholds (Brown and Brown 1996). When this happens, resource availability in a patch may change as a direct response to increasing exploitation of the resource. For example, large seabird colonies seriously deplete local waters, creating a "halo" of unproductive foraging areas (Ashmole 1963; Cairns 1992). This halo develops with time: sites that may have started the season with abundant local food resources may show significant food depletion as a consequence of the activities of the many colony residents. Resource measurements at these times might reveal less food resource per individual in habitat patches with large colonies than in those supporting small colonies, although initial settlement may have been based on the reverse pattern. This illustrates that measuring resource availability to test the ideal free distribution for breeding colonies must take into account time of year, the time when birds choose colonies, and the nature of any group-size effect on resource use. The social costs and benefits associated with different colony sizes and their consequences thus must be known before colony size as an ideal free distribution can be properly evaluated.
Given these difficulties and the fact that only some of the costs and benefits of group size have been identified for most colonial species, it is not surprising that we lack solid evidence that colony size reflects an ideal free distribution of birds. This should not deter investigation, however, especially when resources can be measured directly (e.g., Gibbs et al. 1987; Gibbs and Kinkel 1997). It seems likely to us that heterogeneity in resources within a habitat contributes at least in part to colony-size variation among birds in general, at least in the colony selection phase and even in species in which the social costs and benefits of group size have a large separate influence on fitness. An ideal free process is central to recent models for colony formation (Danchin and Wagner 1997), and for this reason we need field studies specifically designed to measure colony resources. Colony-size variation allows us to make specific predictions about what such studies should find.
We emphasise that measuring habitat resources tells us something very different from measuring reproductive success as advocated by Danchin and Wagner (1997). Reproductive success at a site reflects both the quality and quantity of local resources and the net effect of the social costs and benefits of group size. Thus, knowing reproductive success per se does not reveal the cause of any reproductive differences among colonies and cannot identify whether colonies form mostly in response to favourable habitat features (local resources), for social reasons such as better predator avoidance or food-finding, or both. Measurement of resource availability around colonies of different sizes, coupled with knowing how the social costs and benefits of group size vary, should yield clues to the evolution of coloniality. The extensive development of ideal free theory (reviewed in Tregenza 1995) provides a rich framework for studying variation in colony size.
PHENOTYPIC DIFFERENCES AMONG INDIVIDUALS
Much of the early work on avian coloniality focused on the average expectations of birds in different colonies, commonly analysing how mean values of a cost or benefit varied with colony size. Only recently have differences among individuals within groups been emphasised (Jones 1987; Brown and Brown 1996). Breeding colonies are usually made up of individuals of different ages, with different backgrounds and experience, and of varying physiological condition. These birds may receive different net advantages from colonial nesting as a result of their different attributes; for example, the greater overall vigilance inherent in a large colony may improve the odds that a naive, young bird will survive the breeding season more so than for an older, experienced individual. The implication is that individuals of different abilities may be joining colonies for different reasons. This obviously complicates the analysis of coloniality's origins but probably forces a more realistic emphasis on individual decisions.
Colony-size variation may reflect the different expectations of individuals. Among foraging animals, competitive ability may affect choice of group size: low-ranking fish prefer to feed solitarily rather than in a group because they are poor at competing for food in large groups (Ranta et al. 1993). The notion of sorting among groups based on competitive ability has been applied to groups of cooperatively breeding birds (Brown 1982), fish schools (Ranta and Lindstrom 1990, Ranta et al. 1993), and bird leks (Hoglund et al. 1993). That breeding colonies might reflect sorting of individuals based on personal attributes was suggested by Brown and Brown (1996). Ideal free theory that takes into account unequal abilities of individuals (Sutherland and Parker 1985, 1992; Parker and Sutherland 1986; Jones 1987; reviewed in Tregenza 1995) makes essentially the same prediction that phenotypic characteristics of birds will influence their expectation of success in different habitat patches. We suggest that application of unequal-competitor ideal free models to avian coloniality would be instructive (and see Jones 1987). Colony-size variation provides a natural framework in which to examine phenotypic differences.
We might first ask to what degree colonies differ in their composition. For example, are young, inexperienced, or otherwise inferior individuals relegated to small colonies (as in fish schools), while high-quality birds settle in the larger colonies? If so, this implies that sites differ in their intrinsic quality and that large colonies probably form in the best areas. Small colonies of poor-quality birds may be scattered throughout the habitat in less favourable sites, as occurs for example in Yellow-rumped Caciques Cacicus cela (Robinson 1985, 1986). There are two consequences of such a pattern. It suggests that coloniality evolves through aggregation in areas of high resource availability and is perhaps best explained by an ideal free distribution modified to account for individual differences in competitive ability (reviewed in Tregenza 1995). It also means that observed differences in reproductive success may not be attributed solely to site quality; the characteristics of the birds occupying the site must also be considered. For example, higher reproductive success in larger colonies may be due in part to the features of that habitat patch (local resources) and in part to the social benefits (predator avoidance, food-finding) that accrue whenever large numbers of high-quality birds aggregate.
To date few field studies have examined differences in composition among colony sizes. In Black Skimmers Rynchops niger and perhaps other seabirds, younger birds are suspected to settle in small colonies (Kharitonov and Siegel-Causey 1988; Burger and Gochfeld 1990). In Bearded Tits Panurus biarmicus, females living in colonies tended to weigh more and were thus in better condition than birds nesting as solitaries (Hoi and Hoi-Leitner 1997). If these condition differences applied to the birds at the time they settled and were not merely consequences of group living, Hoi and Hoi-Leitner’s results imply that better quality females aggregate, possibly in areas of high resource abundance. (No differences were found among males that were colonial versus solitary.) In Bank Swallows Riparia riparia, older individuals secured the best nesting sites within colonies, forcing formation of smaller subcolonies composed of younger birds (Jones 1987). Sorting may be passive in which low-quality birds simply avoid the areas of high population density, or it may be mediated by aggressive interactions in which the low-quality birds fail to secure breeding sites in favoured areas by losing contests.
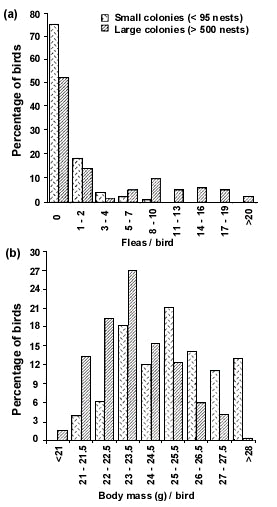
A different situation seems to apply in Cliff Swallows. Age tends to be inversely related to colony size, with birds tending to begin their breeding careers in large colonies and switching to smaller colonies as they get older (Brown and Brown 1996). A possible interpretation is that naive first-year swallows gain more from the antipredator and foraging advantages of a large colony (despite the costs) than do more experienced birds. Older individuals switch to smaller colonies to avoid the substantial costs of larger colonies (ectoparasitism in particular) which apply to all birds regardless of their age. Similarly, birds which are initially parasite free and in relatively good body condition at the time of colony settlement tend to favour smaller colonies (Fig. 2; Brown and Brown 1996). This suggests that high-quality birds who have less to gain from the advantages of a large colony prefer smaller colonies to minimise the costs of group life. Sorting by phenotype illustrates that individuals benefit differently from colonial nesting and suggests that colony size may not always reflect local resource availability or habitat quality. Determining the composition of colonies and the attributes of colony members should thus be a priority in future field studies. Interpreting patterns of reproductive success (Boulinier and Danchin 1997; Danchin and Wagner 1997) will also be influenced by colony composition: how much of the success at a site is determined by intrinsic habitat features and how much by the phenotypic distribution of colony residents? If the latter is a major component of the observed reproductive success, prospective residents are unlikely to learn much about future prospects at a site by observing nesting activity (sensu Danchin and Wagner 1997), unless colony composition between years stays the same.
STABILITY OF COLONY SIZE
Colony size is often considered unstable because an individual may join or leave a colony and thereby change its size (Sibly 1983; Pulliam and Caraco 1984). If there is a single optimal colony size that in theory provides the highest fitness for members, individuals who seek to join the colony from the outside will alter the payoffs for all residents by increasing the colony size above the optimum. Optimal group size thus can be unstable, and groups larger than the optimum might be common (Pulliam and Caraco 1984). Theoreticians have modelled the conditions under which individuals should and should not join groups of particular sizes (Sibly 1983; Giraldeau and Gillis 1985; Giraldeau and Caraco 1993; Higashi and Yamamura 1993; Rannala and Brown 1994). The presumed instability of colony size has led to questions about whether it provides an appropriate framework for analysing fitness of colonial animals (Møller 1997; Danchin and Boulinier in press) and thus whether animals can use it as a reliable cue in selecting where to nest (Kharitonov and Siegel-Causey 1988; Brown and Brown 1996). Others have suggested that site characteristics per se are more likely than colony size to determine fitness (Danchin and Wagner 1997).
We respond to this view in the following ways. Stability of group size depends heavily on the shape of the fitness function associated with group size. If fitness declines steeply in groups larger than the optimal, an individual seeking to join an existing group at optimal size may do better by remaining as a solitary or joining a smaller group (Giraldeau and Gillis 1985). This may occur especially in cases where competition is increased in large groups. Stability of group size will also depend on relatedness between the potential joiner and members in the group: if relatedness is high, a potential joiner may be selected to avoid the group because its presence would depress fitness of all its relatives within the group (Giraldeau and Caraco 1993; Higashi and Yamamura 1993). This probably applies more often to social insects but may also be important for co-operatively breeding birds that live in family groups of various sizes (e.g., Brown 1982). Thus, before one can claim that an optimal colony size is stable or unstable, information on fitness (reproductive success) and relatedness in relation to colony size must be obtained.
The assumption that optimal group sizes exist at all may be problematic, because there is little evidence for optimal breeding colony sizes in general (Brown and Brown 1996). If colonies form in an ideal free way with respect to habitat quality, one would predict no optimal colony size. The lack of a relationship between reproductive success and colony size in the 16 studies mentioned earlier are consistent with there being no optimal colony size, and only seven of 47 studies reviewed in Brown and Brown (1996) provided even weak evidence for an intermediate optimal colony size.
Finally, colony sizes can be stable if individuals co-ordinate their selection of colony site. If individuals seeking to join colonies aggregate prior to settlement (e.g., during migration or foraging), they can colonise entirely new sites and come much closer to achieving a desired colony size than if they each joined an existing group separately (Brown and Brown 1996). In Cliff Swallows, birds move in and out of colonies early in the season in groups, suggesting some collective decision-making. Many sites are first occupied by groups, and birds arrive on the breeding grounds in flocks. Other species show similar mass arrivals and departures at sites (Kharitonov and Siegel-Causey 1988). The assumption that birds looking for colonies operate independently needs to be tested, because this may not be true in general. Colonial birds–well known for their aggregative tendencies–probably show greater parallels with the collective decision-making of the social insects (e.g., Seeley 1995) than generally appreciated. Clearly more must be learned about the mechanisms of colony choice before we can know whether the behaviour of individuals seeking to join groups from the outside represents a potential constraint on achieving a given colony size at a site.
PROSPECTUSWe agree entirely with Danchin and Wagner (1997) that the evolution of coloniality is an evolutionary enigma. None of the current hypotheses are adequate in our view to explain the phenomenon in general, and even well-studied cases (e.g., Brown and Brown 1996) still present many questions and controversies. Colony-size variation may yield important clues to this puzzle. We urge that field workers de-emphasise the classical paradigm of measuring particular costs and benefits in relation to group size and instead concentrate on what the distribution of animals among colony sizes may tell us. If colony-size variation reflects an ideal free match to resources, colonies may form in response to resource heterogeneity in ways similar to foraging groups. If colony composition differs among large and small colonies, individuals may be maximising their fitness in groups of different sizes based on their phenotype and the social costs and benefits of group size. In these cases, we would want to determine how individuals of different phenotypes benefit differently from various group sizes, perhaps through experimental manipulation of colony size after birds have settled.
ACKNOWLEDGMENTS
We thank the 43 research assistants who have helped in our studies of Cliff Swallow coloniality since 1982; the School of Biological Sciences of the University of Nebraska–Lincoln for use of the facilities of the Cedar Point Biological Station during field work; James Gibbs for allowing reproduction of Fig. 1; and (most recently) the National Science Foundation (BSR-9015734, DEB-9224949, DEB-9496113, DEB-9613638), the Erna and Victor Hasselblad Foundation, the National Geographic Society (4545-91, 5469-95), and the University of Tulsa for financial support. For helpful discussion and comments on the manuscript, we are grateful to Etienne Danchin, John Hoogland, and Christine Sas.
REFERENCES
Alexander, R.D. 1974. The evolution of social behavior. Annual Review of Ecology Systematics 5: 325-383.
Ashmole, N.P. 1963. The regulation of numbers of tropical oceanic birds. Ibis 103b: 458-473.
Birkhead, T.R. 1985. Coloniality and social behaviour in the Atlantic alcidae. In: Nettleship, D. N. and Birkhead, T. R. (eds). The Atlantic Alcidae. London; Academic Press: 355-382.
Boulinier, T. & Danchin, E. 1997. The use of conspecific reproductive success for breeding patch selection in terrestrial migratory species. Evolutionary. Ecology 11: 505-517.
Boulinier, T., Danchin, E., Monnat, J.-Y., Doutrelant, C. & Cadiou, B. 1996. Timing of prospecting and the value of information in a colonial breeding bird. Journal of Avian Biology 27: 252-256.
Brown, C.R., & Brown, M.B. 1986. Ectoparasitism as a cost of coloniality in Cliff Swallows (Hirundo pyrrhonota). Ecology 67: 1206-1218.
Brown, C.R. & Brown, M.B. 1987. Group-living in Cliff Swallows as an advantage in avoiding predators. Behavioral Ecology and Sociobiology 21: 97-107.
Brown, C.R. & Brown, M.B. 1996. Coloniality in the Cliff Swallow: The Effect of Group Size on Social Behavior. Chicago; University Chicago Press.
Brown, C.R. & Rannala, B. 1995. Colony choice in birds: models based on temporally invariant site quality. Behavioral Ecology and Sociobiology 36: 221-228.
Brown, C.R., Stutchbury, B.J. & Walsh, P.D. 1990. Choice of colony size in birds. Trends in Ecology and Evolution 5: 398-403.
Brown, J.L. 1982. Optimal group size in territorial animals. Journal of Theorical Biology 95: 793-810.
Burger, J. 1981. A model for the evolution of mixed-species colonies of ciconiiformes. Quarterly Review of Biology 56: 143-167.
Burger, J. & Gochfeld, M. 1990. The Black Skimmer: Social Dynamics of a Colonial Species. New York; Columbia University Press.
Burger, J. & Gochfeld, M. 1991. The Common Tern: Its Breeding Biology and Social Behavior. New York; Columbia University Press.
Cairns, D.K. 1992. Population regulation of seabird colonies. In:. Power, D. M. (ed). Current Ornithology. Vol. 9; New York; Plenum: 37-61.
Crook, J.H. 1965. The adaptive significance of avian social organizations. Symposium of the Zoological Society of London 14: 181-218.
Danchin, E. & Wagner, R.H. 1997. The evolution of coloniality: the emergence of new perspectives. Trends in Ecology and Evolution 12: 342-347.
Danchin, E. & Boulinier, T. In press. Breeding habitat selection based on conspecific reproductive success: the performance-based conspecific attraction hypothesis for the evolution of coloniality. Colonial Waterbirds.
Fretwell, S.D. & Lucas, H.L., Jr. 1970. On territorial behavior and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheoretica 19: 1-36.
Gibbs, J.P. & Kinkel, L.K. 1997. Determinants of the size and location of Great Blue Heron colonies. Colonial Waterbirds 20: 1-7.
Gibbs, J.P., Woodward, S., Hunter, M.L. & Hutchinson, A.E. 1987. Determinants of Great Blue Heron colony distribution in coastal Maine. Auk 104: 38-47.
Giraldeau, L.-A. & Gillis, D. 1985. Optimal group size can be stable: a reply to Sibly. Animal Behaviour 33: 666-667.
Giraldeau, L.-A. & Caraco, T. 1993. Genetic relatedness and group size in an aggregation economy. Evolutionary Ecology 7: 429-438.
Higashi, M. & Yamamura, N. 1993. What determines animal group size? Insider-outsider conflict and its resolution. American Naturalist 142: 553-563.
Hoglund, J., Montgomerie, R. & Widemo, F. 1993. Costs and consequences of variation in the size of ruff leks. Behavioral Ecology and Sociobiology 32: 31-39.
Hoi, H. & Hoi-Leitner, M. 1997. An alternative route to coloniality in the Bearded Tit: females pursue extra-pair fertilizations. Behavioral Ecology 8: 113-119.
Hoogland, J.L. & Sherman, P.W. 1976. Advantages and disadvantages of Bank Swallow (Riparia riparia) coloniality. Ecological Monographs 46: 33-58.
Horn, H.S. 1968. The adaptive significance of colonial nesting in Brewer's Blackbird (Euphages cyanocephalus). Ecology 49: 682-694.
Houston, A.I. & McNamara, J.M. 1988. The ideal free distribution when competitive abilities differ: an approach based on statistical mechanics. Animal Behaviour 36: 166-174.
Hunt, G.L., Jr. & Schneider, D.C. 1987. Scale-dependent processes in the physical and biological environment of marine birds. In: Croxall, J. P. (ed). Seabirds, Feeding Ecology and Role in Marine Ecosystems. Cambridge; Cambridge University Press: 7-41.
Jones, G. 1987. Colonization patterns in sand martins Riparia riparia. Bird Study 34: 20-25.
Kharitonov, S.P. & Siegel-Causey, D. 1988. Colony formation in seabirds. In: Johnston, R. F. (ed). Current Ornithology. Vol. 5; New York.; Plenum: 223-272.
Milinski, M. 1988. Games fish play: making decisions as a social forager. Trends in Ecology and Evolution 3: 325-330.
Møller, A.P. 1987. Advantages and disadvantages of coloniality in the swallow, Hirundo rustica. Animal Behaviour 35: 819-832.
Møller, A.P. 1997. Review of Coloniality in the Cliff Swallow. Ethology 103: 437-438.
Parker, G.A. & Sutherland, W.J. 1986. Ideal free distributions when individuals differ in competitive ability: phenotype-limited ideal free models. Animal Behaviour 34: 1222-1242.
Pulliam, H.R. & Caraco, T. 1984. Living in groups: is there an optimal group size? In: Krebs, J. R. and Davies, N. B. (eds). Behavioural Ecology. Sunderlund, Mass.; Sinauer: 122-147.
Rannala, B.H. & Brown, C.R. 1994. Relatedness and conflict over optimal group size. Trends in Ecology and Evolution 9: 117-119.
Ranta, E. & Lindstrom, K. 1990. Assortative schooling in three-spined sticklebacks? Annales Zoologici Fennici 27: 67-75.
Ranta, E., Rita, H. & Lindstrom, K. 1993. Competition versus cooperation: success of individuals foraging alone and in groups. American Naturalist 142: 42-58.
Robinson, S.K. 1985. Coloniality in the Yellow-rumped Cacique as a defense against nest predators. Auk 102: 506-519.
Robinson, S.K. 1986. Competitive and mutualistic interactions among females in a neotropical oriole. Animal Behaviour 34: 113-122.
Seeley, T.D. 1995. The Wisdom of the Hive. Cambridge, Mass.; Harvard University Press.
Shields, W.M. & Crook, J.R. 1987. Barn Swallow coloniality: a net cost for group breeding in the Adirondacks? Ecology 68: 1373-1386.
Shields, W.M., Crook, J.R., Hebblethwaite, M.L. & Wiles-Ehmann, S.S. 1988. Ideal free coloniality in the swallows. In:. Slobodchikoff, C.N. (ed.). The Ecology of Social Behavior. San Diego; Academic Press: 189-228.
Sibly, R.M. 1983. Optimal group size is unstable. Animal Behaviour 31: 947-948.
Siegel-Causey, D. & Kharitonov, S.P. 1990. The evolution of coloniality. In:. Power D.M. (ed.). Current Ornithology. Vol. 7; New York; Plenum: 285-330.
Sutherland, W.J. & Parker, G.A. 1985. Distribution of unequal competitors. In: Sibly, R.M. and Smith, R.H. (eds). Behavioural Ecology: Ecological Consequences of Adaptive Behaviour. Oxford; Blackwell: 255-274.
Sutherland, W.J. & Parker, G.A. 1992. The relationship between continuous input and interference models of ideal free distributions with unequal competitors. Animal Behaviour 44: 345-355.
Tregenza, T. 1995. Building on the ideal free distribution. Advances in Ecological Research 26: 253-307.
Van Vessem, J. & Draulans, D. 1986. The adaptive significance of colonial breeding in the Grey Heron Ardea cinera: inter- and intra-colony variability in breeding success. Ornis Scandinavica 17: 356-362.
Veen, J. 1977. Functional and causal aspects of nest distribution in colonies of the Sandwich Tern (Sterna s. sandvicensis Lath.). Behaviour Supplement 20: 1-193.
Wiklund, C.G. & Andersson, M. 1994. Natural selection of colony size in a passerine bird. Journal of Animal Ecology 63: 765-774.
Wittenberger, J.F. & Hunt, G.L., Jr. 1985. The adaptive significance of coloniality in birds. In: Farner, D. S. and King, J. R. (eds). Avian Biology. Vol. 8; San Diego; Academic Press: 1-78.
Fig. 1. Great Blue Heron colony size in relation to availability of tidal wetlands (feeding areas) within the colony’s foraging range in coastal Maine, USA. Colony size increased significantly with availability of wetlands (r = 0.79). From Gibbs et al. (1987).
Fig. 2. Percentage distributions of Cliff Swallows carrying various numbers of fleas (a) and of different body masses (in g; b) at the time of settlement in small colonies and large colonies early in the spring in 1988 in southwestern Nebraska, USA. The distributions for small and large colonies differed significantly for fleas per bird (c28 = 45.2, P < 0.001) and for body mass per bird (c28 = 58.7, P < 0.001). Sample sizes were 132 birds for small colonies and 206 birds for large colonies. From Brown and Brown (1996).