S18.5: Understanding risks to birds from pesticide-treated seeds
Andrew Hart, Steven Fryday, Helen McKay, Juan Pascual & Philip Prosser
Central Science Laboratory, Sand Hutton, York, YO41 ILZ, UK, fax 44 1904 462111, e-mail a.hart@csl.gov.uk
Hart, A., Fryday, S., McKay, H., Pascual, J. & Prosser, P. 1999. Understanding risks to birds from pesticide-treated seeds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1070-1087. Johannesburg: BirdLife South Africa.Applying pesticides to seeds before sowing protects crops, but presents a potential risk to seed-eating birds. Using field and laboratory studies we have examined a range of factors which determine whether birds will be poisoned by treated seeds. These include the area sown, the toxicity of the pesticide, its concentration on the seed, the density of exposed seed, the availability of other foods and the ability of birds to selectively avoid treated seed. Avoidance was shown to be complex. Its effectiveness in preventing poisoning depends crucially on whether birds stop feeding before ingesting a lethal dose. This depends on feeding rate, which in turn depends on several factors including hunger, daily feeding patterns and group size. Even when a lethal dose is ingested, some birds regurgitate enough to survive. Taken together these factors imply that some seed treatments used in the UK will poison birds but only occasionally, under particular circumstances. Three sources of evidence including radio-tracking confirm that the actual frequency of poisoning is consistent with this assessment. These findings provide new insights into the responses of birds to contaminated foods, and have important implications for the methods used to predict, monitor and reduce risks to birds from pesticides.
INTRODUCTION
Seed treatments are often promoted as an efficient, environmentally friendly, low-input use of pesticides. One disadvantage of seed treatments, however, is that they may present a high risk to seed-eating birds (Murton & Visozo 1963; Stanley & Bunyan 1979).
A few of the insecticidal seed treatments used in the UK are sufficiently toxic that birds feeding exclusively on treated seed would be expected to ingest several lethal doses per day. Investigations of wildlife mortality incidents show that these seed treatments do cause poisoning (see below) but less often than might be expected from the worst-case risk assessment. This could be due to over-estimation of risk, and/or under-reporting of incidents. There was therefore a need to investigate the relationship between predicted and actual risks to birds from seed treatments, and if necessary develop improved methods for risk assessment.
Some seed treatments can be shown to be repellent to captive birds, and such studies are increasingly used by pesticide manufacturers to argue that exposure will be limited in the wild. Prior to this study it had yet to be demonstrated that low palatability in the laboratory is a reliable indicator that a formulation will be avoided in the wild. There was therefore a need to test whether avoidance could be detected in the wild, and identify the types of laboratory test that are most reliable as a basis for predicting it.
This project was initiated to address these issues through a combination of field and laboratory studies. Two of the main objectives were:
(1) to measure the actual risk to birds from seed treatments, and
(2) to identify factors affecting exposure of birds to seed treatments.
It is important to emphasise that this project focussed almost exclusively on the risk of mortality from short-term exposures, with the aim of ensuring this could be reliably predicted for regulatory purposes. Some of the data are also relevant to assessing longer-term exposure and sub-lethal effects, but these are not addressed in this paper.
The aim of this paper is to illustrate the understanding of risks from treated seeds which has developed in the course of this project. It provides an overview of results from an extensive series of field and laboratory studies carried out over several years. Some of the detailed results are published elsewhere (as cited below), and other publications are in preparation. The paper concludes with a brief discussion of some of the implications for risk assessment and risk management.
METHODS
Species and pesticides studied
In order to obtain the detailed understanding required to address the project objectives it was necessary to focus most of the field work on one pesticide and one species of bird. Woodpigeons Columba palumbus and the insecticide fonofos (O-ethyl S-phenyl (RS)-ethylphosphonodithioate) were chosen. It is important to stress that these were used solely as experimental models for other species and seed treatments.
Fonofos was chosen because it was currently the most widely used of the insecticidal seed treatments with high avian toxicity (it has since been withdrawn from sale in the UK by the manufacturers). However, the method used to measure mortality in the wild provided for the detection of the poisoning by any organophosphorus pesticide. Woodpigeons were chosen because they are one of the species most commonly involved in poisoning incidents caused by seed treatments. They are large enough to carry radio-transmitters with a battery life of several months, they are relatively easy to observe feeding, and their general ecology is well known from earlier studies.
Woodpigeons are very wary of humans and therefore impractical for intensive study in captivity. Most of the laboratory studies were therefore conducted with Feral Pigeons Columba livia, which are closely related to Woodpigeons and have a similar diet. The broad comparability of the two species in their response to fonofos was confirmed experimentally during the project (see later).
Overview of methods
Study sites
The field studies were conducted in fenland near Cambridge, England, in an intensive agricultural area with a large Woodpigeon population, where fonofos was used relatively widely to control Wheat Bulb Fly Delia coarctata. Experiments with captive birds were conducted at our laboratories, except for the pen study (see below).
Measuring mortality of Woodpigeons in the wild
Woodpigeons were captured and fitted with radio-transmitters, then released. The transmitters were temperature-sensitive so that the signal changed when birds cooled down after death. Signals were monitored daily during the winter sowing period and an immediate search was made for any suspected mortalities by following the signal. Post-mortem examination and residue analysis were used to determine the cause of death of those birds which were recovered. The results were used to estimate the proportion of the population dying from poisoning by seed treatments.
Feeding preferences and avoidance of treated seed
Each winter a large number of fields on a circular route were visited at least once per week. On each visit the number of Woodpigeons present on each field was recorded. The results were used to assess the preferences of Woodpigeons for cereals and other crops, and to test whether they avoided fields sown with fonofos-treated wheat. Other methods were also used to test for avoidance: continuous observation of treated and untreated cereal fields for several days after sowing, and monitoring the density of exposed seed to test whether fonofos-treated seed disappeared more slowly.
Other field studies
Data were collected on the densities of seeds exposed on cereal fields after sowing, and on the levels of fonofos they contained at different times after sowing. Dead Woodpigeons were obtained from local shooters and dissected to assess the proportion which had fed on cereal seed and the level of fonofos residues in their gut contents. In one year, video recordings were made at simulated spills of untreated wheat seed, to monitor the range of species which fed on them.
Testing avoidance with captive birds
Most experiments were conducted with Feral Pigeons housed singly or in small groups in outdoor aviaries measuring approximately 3 m x 2 m x 2 m (e.g. Fryday et al., 1998) though some experiments were conducted with singly-caged birds indoors (Pascual, Hart & Fryday, in press). After a period of acclimatisation and preparation (the conditions of which proved to be critical, see later) they were offered wheat seed treated with fonofos, usually at the concentration which was approved in the UK for controlling wheat bulb fly (1080 mg kg-1). In most experiments treated food was offerred without any untreated alternative being available. Experiments lasted 1-2 days during which food consumption, signs of intoxication and mortality were recorded. Additional measurements were made in some experiments including residues of fonofos in the food and crop contents, enzyme activity (cholinesterase) in the brain after death, and video recordings of the birds’ behaviour.
Pen trial
A pen trial was conducted in which Feral Pigeons were held captive for 2 days in pens on field plots sown with fonofos-treated wheat (Pascual and Hart 1997). Observations were made for signs of intoxication, mortality and changes in body weight, and blood samples taken before and after the test were analysed for enzyme activity.
Model
A computer model was devised to simulate the level of mortality which would occur if fonofos were not avoided.
Experiments on sowing depth
Agricultural experiments were conducted to test the effects of sowing depth and post-sowing cultivations (rolling and harrowing) on the density of exposed seed, and hence their availability to birds (Pascual, Hart, Saunders, Mackay, Kilpatrick & Prosser, in press; Pascual, Saunders, Hart & Mottram, in press).
RESULTS AND DISCUSSION
The Actual Risk To Birds From Seed Treatments
Obtaining a reliable measure of the actual risk to birds from seed treatments under current conditions was an essential component of this project. Without this there would be no means of assessing the accuracy of current methods of assessing risk, or evaluating alternatives. Two important measures already existed: poisoning incidents reported to the UK Wildlife Incident Investigation Scheme (e.g. Fletcher et al. 1997), and surveys of Woodpigeons conducted in the same area as our field studies since 1983 by Dr A S Cooke of English Nature (Cooke 1988). However, both of these measures are likely to under-estimate the true frequency of affected birds, because of under-reporting and under-recording respectively. In addition, the surveys recorded sublethally-affected birds and their eventual fate (death or recovery) is unknown. In this study therefore we monitored the survival of radio-tagged Woodpigeons in the Cambridgeshire study area, to obtain a direct estimate of the frequency of poisoning.
A total of 106 birds were radio-tagged in the winters of 1994-95 and 1995-96, but no definite instances of poisoning were detected (Prosser et al. unpublished manuscript). Seven birds were returned by hunters after being shot. Five birds were found dead and largely intact. The crop contents of one of these contained residues of the insecticide fonofos but at a level (4 mg kg-1) unlikely to have caused death. This carcass was severely scavenged, consisting of little more than the crop, bones and small shreds of other tissues, so alternative causes of death cannot be identified; it also smelled strongly of fox urine, which could result either from death by predation or from scavenging after death by another cause. In 10 cases insufficient tissue was recovered for analysis. A definite diagnosis is not possible for these birds: they may have been taken by predators, or eaten by scavengers after being shot or poisoned. Taking account of these uncertainties, it is concluded from the radio-tracking results that the actual probability of poisoning per Woodpigeon per winter for the local population was unlikely to exceed 5%, and was probably close to zero.
This estimate indicates that the actual risk to Woodpigeons from OP seed treatments is relatively low, and that its influence on population levels is much smaller than that of hunting or changes in cropping patterns (Inglis et al.1997). The estimate is consistent with the other two measures for the same period, which also indicate a low frequency of poisoning (Prosser et al. unpublished manuscript). Only 3 incidents involving OP seed treatments and birds were reported to the Wildlife Incident Investigation Scheme (WIIS) in 1994-1996, one of which involved Woodpigeons. Of over 20,000 Woodpigeons counted in Dr Cooke’s surveys in the same winters, none were recorded as being affected.
WIIS and the surveys also indicate a similarly low incidence of effects in the early 1990s. However, both suggest a significantly higher incidence of effects in the 1980s, concentrated mainly in three particular winters. WIIS recorded a total of nine Woodpigeon incidents in 9 years, and the frequency of sublethal effects observed in roost surveys averaged 6% of roosting Woodpigeons per winter (16% in the worst winter). This may reflect the higher usage of OP seed treatments in the early 1980s, and/or the replacement of chlorfenvinphos by fonofos (which may be less persistent) in the 1990s, and/or changing agricultural practice (e.g. improved incorporation of seed).
It is important to note that all these results are specific to Woodpigeons: the risks to other species may be higher or lower. Feral and racing pigeons Columba livia, Grey Partridge Perdix perdix and Ring-necked Pheasant Phasianus colchicus have also been recorded in WIIS incidents since 1981, and other species may have been affected but not reported. Differences between species are considered further below.
In summary, the results show that the frequency of poisoning of Woodpigeons by OP seed treatments was low and probably close to zero in the 1990s, but reached appreciable levels in some years in the 1980s.
Factors affecting the exposure of birds to seed treatments
Pesticide usage
Variation occurs in the area and timing of pesticide usage, influencing both the number of birds potentially exposed and the degree of exposure of individual birds. Data from the Pesticide Usage Survey (e.g. Garthwaite et al. 1995) show that the total usage of OP seed treatments on cereals in the UK fell from around 150,000 ha to 40,000 ha between 1982 and 1988, then remained steady at 40-50,000 ha in 1990, 1992 and 1994. There was also a substantial, and perhaps ecotoxicologically significant, shift in the balance of active substances, with chlorfenvinphos dominating in 1982 and fonofos from 1988 onwards. In our Cambridgeshire study area in the years 1992-1996, about half the cereals sown were treated with fonofos. Most of the fonofos-treated fields were sown between mid-October and mid-December, but each year a few were sown in the Spring (January - April). Most other cereal fields were sown with seed treated only with fungicides: chlorfenvinphos was rarely used. The spread of sowing dates meant that in any week, only a few fields contained newly-sown fonofos-treated wheat.
This pattern of usage would lead one to expect a decline in risk during the 1980s, and an annual peak in October-December. This agrees well with the actual trends, both in the frequency of poisoning incidents reported to WIIS and in the frequency of sublethally-affected Woodpigeons recorded in surveys at Woodwalton Fen (Cooke 1988).
Residue loss
The nominal concentration of fonofos on treated wheat is 1080 mg kg-1, but in our samples concentrations were always lower than this. Seed taken from the drill hopper, or treated in small batches for aviary experiments, typically contained 600-800 mg kg-1. A decrease of about 65% occurred within a day, possibly by abrasion during the sowing operation itself, so that the concentration on exposed seed on the day after sowing averaged 241 mg kg-1 in 1994 (McKay et al. in press). After this the rate of decay was much lower, about 2% per day. Similar results were obtained in earlier years. It was also shown that decreases of similar order but over a longer period can occur when seed is stored for periods of over a month as may happen if farmers are prevented by adverse weather from sowing when they intend.
These decreases in residues compared to the nominal concentration clearly have a significant impact on risk for fonofos. The rate of residue loss was not examined for other seed treatments, but may merit increased attention in risk assessments.
Density of exposed seed
High densities of seed exposed on the soil surface may increase exposure and hence risk in two ways: by attracting more birds to feed on the field, and by increasing the rate at which they can feed.
The density of exposed seed was found to vary over a very wide range, both within and between fields, even though soil type and conditions in the study area generally favour good incorporation of seed (Pascual, Hart, Saunders, Mackay, Kilpatrick & Prosser, in press). Densities were about twice as high in headlands (average 10 seeds m-2) as in the rest of the field: this will be especially significant for those species of birds which feed mostly at the field edge. Density was related to sowing depth: densities over 10 seeds/m-2 were found mostly where the actual sowing depth was less than about 40 mm (Pascual, Hart, Saunders, Mackay, Kilpatrick & Prosser, in press). Density decreased after sowing at an average rate of about 2% per day (McKay et al. in press). This may be due partly to physical processes (rain, wind, settling), partly to post-sowing cultivations such as rolling and harrowing when these were performed (Pascual, Saunders, Hart & Mottram, in press), and partly to removal of seed by birds and other animals.
Seed densities in other areas may differ from those found in this study due to differences in soil type and conditions and variations in husbandry, but the qualitative results (wide variability, higher densities in headlands, inverse correlation with sowing depth, effects of cultivations) probably apply generally.
Experiments with captive pigeons showed they will dig to obtain seed buried in soil at depths similar to sown seed. However, observations of wild Woodpigeons showed they spent much less time digging than they did pecking at exposed seed, and it is possible that digging is related to the presence of larger buried items such as potato waste. Presumably pigeons can ingest exposed seed at much higher rates than buried seeds, except when the density of exposed seeds is very low. This is likely to apply also to other species. In view of the high feeding rates required to cause lethal poisoning in captive birds (see below) it is concluded that the contribution of buried seeds to risk is negligible.
Frequency of seed spills
Spills of seed are likely to present increased risk for three reasons. First, the density of exposed seed is very high, which will tend to attract birds and enable them to feed at maximum rates. Second, seed which is spilled during transporting or loading may carry higher residues of pesticides than seed which has been sown if losses occur during sowing (see above). Third, spills often contain sufficient seed to poison several individuals. The frequency of seed spills was not recorded in the present study but an average of 1.7 spills per field was found for wheat fields in the Netherlands by de Leeuw et al. (1995).
Attractiveness of newly-sown cereals
Surveys showed that cereal fields were more attractive to Woodpigeons in the first week or so after sowing than they were subsequently (McKay et al. in press). Decreasing attractiveness may be partly due to the density of exposed seed decreasing with time after sowing (see above), though the relationship between seed densities and bird numbers was too variable to confirm this. It could also be due to the depletion of any other food which may be present, such as residues of the previous crop (though this was not measured in this study).
It has been supposed that a general scarcity of food in the spring might make spring wheat more important to birds than winter wheat, and that seed treatments might therefore pose higher risks in spring than autumn. Newly-sown cereals did appear to be attractive to Woodpigeons in spring but, due to the limited occurrence of spring sowings in the study area, a robust comparison with autumn sowings was not possible.
Other foods
The attractiveness of cereal fields will depend partly on what other foods are available, which varies seasonally, and their relative attractiveness to birds. Harvest waste from root crops in winter (mainly potatoes) and the newly-sown seed of some spring-sown crops (peas, beans and onions) were shown to be attractive to Woodpigeons (McKay et al. in press). When these are available they will tend to reduce the extent to which Woodpigeons feed on cereal seed.
Of 61 shot Woodpigeons, only 3 (5%) had been feeding exclusively on cereal seed (McKay et al. in press). A further 43% contained cereal seed together with other foods including other crop seeds (beans, peas, maize and rape), unidentified foliage and natural foods (berries, rose hips, acorns, fungi and snails). However, for those birds which contained it, cereal seed accounted for 47% of the crop contents (by weight). This shows that although most birds are taking a mixed diet, many of them are still ingesting significant amounts of cereal.
Avoidance
It was known before the start of the project that birds may reduce their consumption when feeding on fonofos-treated food, as this occurs with Japanese quail in the standard 5-day dietary study (LC50; see example for fonofos in Hill and Camardese 1986). This ‘avoidance’ response has the potential to limit exposure and risk. A major part of this project was committed to testing whether this response occurs in the wild, and to investigating the factors which affect it.
Surveys showed that, in the first week after sowing, Woodpigeons were less likely to be present on fields sown with fonofos-treated wheat than on other wheat fields (McKay et al. in press). The same result was obtained in 4 successive winters. Fields with the highest residues of fonofos on the seed were the most strongly avoided. These results suggest that the avoidance response does occur in the wild.
The importance of the avoidance response was assessed by considering what the frequency of poisoning by fonofos would be, if birds showed no tendency to avoid it. It is not possible to answer this experimentally because avoidance cannot be ‘switched off’. Two approaches were attempted. First, comparison with historical data for aldrin and heptachlor, which were used as seed treatments in the 1960s. Murton and Visozo (1963) estimated that at least 8% and 'probably very much more' of a local population had been poisoned, during a period of a few days. The level of mortality caused by fonofos is clearly much lower. This is unlikely to be due to differences in toxicity (LD50 or LC50) as when these are compared to the concentrations used on seed, they indicate a greater risk for fonofos than the organochlorines. That mortality is actually lower for fonofos may be partly due to avoidance. It could also be due to other factors such as the accumulative action of the organochlorines and differences between the 1960s and 1990s in the proportion of fields sown with treated cereal and the density of exposed seeds.
The second approach to assessing the importance of the avoidance response was the construction of a quantitative model which included effects of usage, residues, seed density and alternative foods on the frequency of poisoning, but excluded avoidance (simulating the effect of ‘switching off’ avoidance). The results suggest that, if fonofos were not avoided, poisoning would be frequent, and involve large numbers of birds which would be very likely to be reported to the authorities (Hart et al. unpublished manuscript). In fact, poisoning incidents are very infrequent and involve only one or a few birds. This result suggests that avoidance probably does contribute significantly to preventing mortality due to fonofos. However, it must be treated with caution, as the model necessarily made a number of simplifying assumptions which may be causing substantial over- or under-estimation.
A series of experiments were conducted with captive Feral Pigeons to investigate the basis of the avoidance response and why it sometimes fails to prevent mortality (as shown by the occurrence of poisoning incidents). When given a choice between treated and untreated seed in separate pots, birds ate an average of only two treated seeds before learning to avoid the rest and exhibited no signs of intoxication. In all subsequent experiments the birds were offered only treated seed.
It was shown that more treated seed was consumed when it was presented spread on a tray, rather than in a pot (Fryday et al., 1998). This may be due to the effects of pesticide vapour being more concentrated over the pot. As this is unlikely to occur in the wild, trays were used in all subsequent experiments.
The strength of the avoidance response was concentration-dependent. In one experiment, avoidance was detectable at 75 mg kg-1 seed and increased sharply at higher concentrations. At concentrations above 140 mg kg-1 consumption was decreased to less than 20% of normal. This compares well with the concentration above which avoidance was observed in the surveys of wild Woodpigeons (150 mg kg-1). However, further experiments were required to discover under what conditions avoidance breaks down, and explain the occurrence of mortality in the wild (see below). Because of this emphasis on identifying the conditions under which mortality occurs, seed was treated at the approved rate (1080 mg kg-1) in all subsequent experiments, producing measured concentrations of 600-800 mg kg-1.
Hunger and body condition
In the first experiments birds were allowed unrestricted access to untreated food every day, and deprived of food only overnight before testing. When food was withheld for 24 hours before testing, the extra deprivation did not significantly affect consumption of the treated food. When the deprivation period was extended to six days there was a significant increase in the consumption of treated seed compared to non-deprived birds, though substantial avoidance was still apparent. A similar result was obtained when birds were only given access to food for two hours per day, and allowed only 15% of normal consumption for six days before testing (Pascual, Hart & Fryday, in press). These were the first experiments in the project where birds took a lethal dose of fonofos during their first bout of feeding on treated seed. This was important because, in the wild, it is unlikely that a bird will resume feeding on the same field after the avoidance response has set in: it is more likely to switch to another field or another type of food.
The conditions of these experiments were severe but not unrealistic: six days continuous snow cover occurs on average once per year in the Cambridgeshire study area. Birds in the experiment lost only 11% of their weight, whereas in the wild much larger variations in weight occur. Finally, there was no statistically significant difference in the fat content of the deprived and non-deprived birds. Thus these experiments showed that there are some realistic conditions under which hunger decreases the avoidance response enough to cause mortality.
Social effects
Birds were caged singly in all but one of the experiments mentioned above. In the exception, increasing the group size to two did not change consumption in a consistent way. In a later experiment, birds were housed singly or in groups of four and eight. All the birds were deprived of food for one day before testing, and had previously been trained to take all their food in a two hour period each day. On the test day, birds in groups of four and eight consumed (respectively) twice and three times as much treated seed as single birds. These differences were statistically highly significant, and are presumably due to competitive feeding. Mortality also increased with group size, reaching 84% in groups of eight. This experiment showed that group-feeding was more effective in overcoming the avoidance response than the relatively severe food deprivation of the previous experiments, and increased the range of conditions under which mortality might be expected in the wild.
Feeding rate
Mineau et al. (1994) suggest that differences in the feeding rate of wild and captive birds can explain discrepancies between LC50 data and mortalities in the field. We tested this by restricting pigeons’ feeding period to two hours per day. Birds rapidly learned to take all their daily food within the 2 hours and, when offered treated food, ate over 4 times as much as birds which had been allowed to feed continuously (24 h day-1). Restriction of feeding to 2 hours a day is not unrealistic and could occur during bad weather. Also, birds in the wild are often under pressure to feed fast in order to minimise the time for which they are exposed to predators.
Measurements of feeding rate were obtained in this experiment and several of the earlier experiments, described above. When considered together, the results show that as feeding rate increases, so does the amount of seed ingested before avoidance sets in, and so does the extent of mortality (Fig. 1). This result clearly demonstrates the crucial role of feeding rate in determining risk.
It must be remembered that these results relate to very acute (short-term) exposures. In most cases the avoidance response stops feeding within 10 minutes or less, but the rate of feeding is such that some birds have already ingested a lethal dose. There were differences between experiments in the concentration of fonofos on the seed, which probably contributed to the increased mortality at the highest feeding rates. However, the effect of increased feeding rate is crucial and cannot be simulated by increasing the concentration in the feed: in an experiment where the concentration of fonofos was increased to ten times the recommended rate only one of three birds took a lethal dose, and not in the first bout of feeding.
From these results (Fig. 1) it can be seen that the other factors which were shown to overcome avoidance were acting through their effect on feeding rate. Birds which were hungry or feeding competitively, fed faster. Increasing group size from one to four birds had a stronger influence on feeding rate than was obtained after six days starvation, despite the fact that measured fonofos residues were higher in the experiment on group sizes (931 vs. 693 mg kg-1), which would be expected to decrease feeding rate.
Understanding the role of feeding rate also enables us to identify other factors which will influence risk: feeding rate and hence risk should be reduced when the density of exposed seeds is low, or when birds have to dig for buried seeds. Feeding rate should also be reduced if birds have to remove the husk or manipulate each seed in other ways before swallowing it (Avery et al. 1997).The critical question for risk assessment is whether the rate of ingestion is slow enough to allow time for avoidance to set in before a lethal dose is obtained.
These results demonstrate that an understanding of the influence of feeding rate is crucial to predict the risk of mortality from acute exposures to treated seed. Two types of information are required. First, information is needed on the relationship between feeding rate, dose ingested, and mortality, such as the results presented in Fig. 1. In principle, such data could be obtained in a single experiment in which group size, feeding time and hunger are manipulated to make birds feed at different rates. For Feral Pigeons feeding on seed treated with fonofos at 693-931 mg kg-1, mortality occurred at feeding rates above about 2 g minute-1 (Fig. 1). Second, information is needed on the feeding rates which occur in the wild. We obtained this information from observations of Woodpigeons feeding on newly-sown cereal fields. Over 50% of Woodpigeons fed at less than 0.25 g minute-1, well below the level which caused mortality in the aviary experiments. Only about 1% of wild Woodpigeons fed at over 2 g/minute, the level which caused mortality in captive Feral Pigeons.
In fact, the results in Fig. 1 presumably exaggerate the chance of mortality because the aviary experiments were done with fonofos concentrations of 693 - 931 mg kg-1 whereas concentrations in the wild are usually much lower (mean 241 mg kg-1 on the day after sowing). The results therefore suggest that the feeding rate of wild birds will rarely reach the levels required to overcome the avoidance response, and that mortality will consequently be rare. This agrees well with wildlife incident data and radio-tracking studies (see above) which confirm that the frequency of mortality in the wild is very low.
Regurgitation
In the experiment on feeding rate it was observed that two birds regurgitated some of the treated seed they had eaten, and it was recognised that this might significantly reduce the dose absorbed. Consequently, in the experiment where Feral Pigeons were restricted to 15% of normal food consumption for six days before testing, detailed residue analyses were used to quantify the proportions of the initial dose which were absorbed and regurgitated (Pascual, Hart & Fryday, in press). The results showed that under these conditions avoidance alone was insufficient to prevent ingestion of a lethal dose, and regurgitation was crucial in enabling half the birds to survive. However, the extent of regurgitation was variable: two birds did not regurgitate at all, while the remaining six regurgitated amounts ranging from 61 - 98% of the ingested seed. Regurgitation reduced the level of mortality but did not eliminate it, as some birds died without regurgitating and others despite doing so.
Other experiments showed that the extent of regurgitation by Feral Pigeons varied with feeding rate and with the concentration of fonofos on the seed.
These studies demonstrate for the first time that regurgitation can reduce the mortality of birds exposed to pesticides in their food, providing them with a second mechanism of defence against poisoning (the first being the avoidance response). Earlier investigations with emetics show that a wide range of bird species are capable of regurgitation but that the intensity of the response is highly variable within and between species (reviewed in Hart and Thompson 1995). It is concluded that regurgitation can contribute to reducing risk in those species which are capable of it, but may be ineffective under some conditions and may not protect all the individuals which are exposed. This variability makes it difficult to predict the influence of regurgitation in risk assessment. However, it is important to avoid under-estimating risk when using test data for species which can regurgitate, to predict risk for species which cannot (Pascual, Hart & Fryday, in press).
Repeated exposure
Birds might learn to avoid treated seed more effectively as a result of their first exposure to it. An experiment was conducted with Feral Pigeons to test this. The results clearly showed a learned enhancement of avoidance, which persisted at least 4 days after the first exposure (Fryday et al. unpublished manuscript). This learned avoidance should act to reduce risk in the wild. It is even conceivable that individual birds are only at significant risk on their first encounter with treated seed in each sowing period as, within sowing periods, they are likely to encounter treated seed frequently enough to maintain the learned avoidance. However, further work would be required to determine how long the learned enhancement persists if it were desired to quantify its contribution to reducing risk.
Birds leaving a field sown with treated seed may sometimes encounter another treated field on the same day. In this case, pesticide ingested at the first field might not have been metabolised, and the birds might ingest enough additional pesticide at the second field to exceed lethal levels. This possibility was also examined experimentally but the results were inconclusive because, under the conditions used, most birds were too severely affected by the first exposure to feed again within the day (Fryday et al. unpublished manuscript).
Species differences
Due to the nervous nature of Woodpigeons in captivity and therefore the difficulty of working with this species, Feral Pigeons were used as models for Woodpigeons in most experiments. A simple test was therefore conducted to check that there were no gross differences in response to fonofos between the two species (Fryday et al. unpublished manuscript).
The birds were tested in large groups. Both species showed substantial avoidance, consuming less than half their normal daily intake on the test day. In both species the mechanism of avoidance appeared to be post-ingestional, and both species regurgitated some seed. Woodpigeons waited longer to start feeding, and fed for a shorter period before stopping, but fed nearly twice as fast as Feral Pigeons.
Mortality occurred in both species. However, only two of 14 Woodpigeons died, compared to 14 of 16 Feral Pigeons. The intake of fonofos per unit bodyweight was smaller for Woodpigeons than Feral Pigeons. This may account for the difference in mortality, but a difference in toxicity between the two species cannot be ruled out.
These results show that avoidance and regurgitation occur in Woodpigeons as well as Feral Pigeons, and that in both species these mechanisms can fail to prevent lethal poisoning when birds feed in flocks. It is not known how far these findings apply to bird species in general, though it should be noted that pigeons may be unusually adapted to regurgitation as they use it to feed their young.
Risk might be affected by differences between species in the extent to which they feed on cereal seed, and whether they dehusk it (thus removing some of the pesticide coating, Avery et al. 1997). This was investigated by using video cameras to record birds feeding at patches of untreated seed placed at the edge of fields. The most frequent visitors were Robins Erithacus rubecula, Ring-necked Pheasants and Chaffinches Fringilla coelebs, and of these only the Chaffinch was seen to husk seeds. Further work is underway using this method to investigate the species which take other types of agricultural seed.
Environmental factors
There are a number of mechanisms by which weather conditions could influence the risk to birds from poisoning by seed treatments. Possibilities include:
Prolonged or heavy rain may make the soil wet, reducing sowing efficiency or causing a few farmers to broadcast seed in contravention of pesticide label instructions. This would lead to increased densities of exposed seed, attracting more birds and enabling them to feed at a faster rate which increases risk.
Low temperatures or prolonged periods of reduced food availability due to frost or snow may make birds hungrier, causing them to feed faster or over-ride the avoidance response.
Low temperatures may increase the susceptibility of birds to organophosphorus compounds, for example if the pesticide interferes with thermoregulation.
It was decided to look for evidence of these effects by analysing the temporal distribution of past poisoning incidents in relation to the weather. The number of poisoning incidents reported to the Wildlife Incident Investigation Scheme (WIIS) was too low for such an analysis. Instead, data were obtained on the frequency of apparent sublethal poisonings (some of them confirmed by WIIS) recorded during regular surveys of a large Woodpigeon roost by Dr A S Cooke over 14 winters. These data were analysed in relation to local weather records.
None of the possibilities suggested above were strongly supported by the analysis (Hart et al. unpublished manuscript). Most observations of abnormal behaviour were recorded in 3 winters which all followed high rainfall in September/October. However, looking at short-term patterns, the occurrence of abnormal behaviour was significantly associated with periods of lower rainfall. It seems likely that this reflects the tendency of farmers to drill cereals when the soil is dry. The strongest pattern in the data is a decrease in the frequency of abnormal behaviour between the 1980s and the 1990s. As stated earlier, this may reflect changes in the usage of seed treatments in the area. It is concluded that weather conditions are not the primary determinant of risk, but the data are inadequate to detect more subtle effects.
Relative importance of factors affecting risk
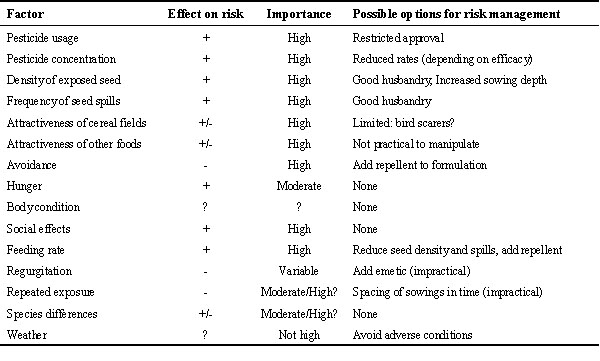
The relative importance of the various factors affecting risk can be assessed qualitatively from the results discussed above (Table 1). The available data are not adequate to assess the importance of all the factors quantitatively, though this is possible for some factors. For example, the experiments with captive pigeons quantified the role of feeding rate and showed that it is more affected by group size than hunger. The influence of body condition and weather remains uncertain, though the results indicate that these are probably less important than most of the other factors which were investigated.
The factors can be divided into three groups: those which determine the availability of treated fields (pesticide usage), those which determine how many birds visit those fields (dietary preferences, seed density, degree of disturbance, social factors), and those which determine the degree of risk during each visit (pesticide concentration, seed density, spills, avoidance, feeding rate, social effects, hunger, regurgitation, previous experience with treated seed, toxicity). This division may be helpful in structuring risk assessment. It is standard practice to concentrate first on what happens when birds are present (i.e. the third group of factors). If it is considered that mortality is likely, then it becomes important to assess how often this may occur (i.e. the first two groups of factors).
The way in which the factors in the third group interact is illustrated in Fig. 2. Three points are worth highlighting here.
The pivotal role of feeding rate and pesticide concentration in determining the speed at which the active substance is ingested.
The fine balance between mortality and survival, depending on whether the net dose reaches the lethal level before the avoidance and regurgitation responses can intervene.
Although potentially both buried and exposed seeds can contribute to the net dose, in practice the time taken to find buried seeds means that they slow the rate of feeding (Pascual, unpublished data) to levels where the avoidance response is fully effective. Consequently the contribution of buried seeds to risk is negligible, and concern and risk management should focus on the density of exposed seeds.
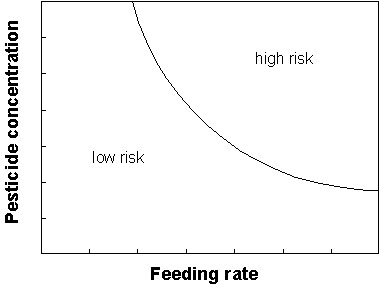
Finally, this project has shown that both pesticide concentration and seed density need to be high in order to generate significant risk. This is illustrated in Fig. 3. The results obtained are not sufficient to determine the position of the boundary between low and high risk. This would be expected to vary between active ingredients, and also between species (due to differences in toxicity, the avoidance response, and the regurgitation response). If a high degree of precision was required in assessing the risk from a particular seed treatment, then it would be necessary to quantify the relationship shown in Fig. 3 for relevant species. This might be done, for example, by conducting experiments with captive birds at a range of combinations of concentration and feeding rate. For active substances with lower degradation rates than fonofos, it might be sufficient to quantify the effect of feeding rate only. It would probably not be necessary to quantify the more detailed factors illustrated in Fig. 2. It would be necessary, however, to consider carefully whether results for one species could reliably be extrapolated to others which might be at risk, since it will not be practical to test all of them.
Risk management
If the factors which are identified as influencing risk can be manipulated they may be suitable options for risk management. This project has identified many such factors (see Table 1), of which the most practical to manipulate are probably the density of exposed seed and the frequency of seed spills. Both these operate by affecting the rate at which birds can feed on treated seed.
The first conclusion is that good agricultural practice should favour good seed incorporation and hence contribute to reducing risk. This includes both good sowing practice and the removal or burying of spillages, both of which were specifically promoted in the UK by the User Education Programme for fonofos which was introduced in 1994.
Results from this project can be used to examine the effectiveness of these risk management options in more detail. Increasing sowing depth was shown to cause substantial reductions in the density of seed exposed on the soil surface, and a high degree of incorporation can be achieved by sowing depths of about 40 mm. This depth was well within the range currently used by farmers in our study area and, in experimental plots, did not cause any decrease in yield compared to sowing at 20 mm (Pascual, Hart, Saunders, McKay, Kilpatrick & Prosser, in press). Additional data on current sowing practices in the UK was obtained in conjunction with the 1996 survey of pesticide usage on arable farms (Garthwaite, pers. comm.). The results showed a wide variation in sowing depths reported by farmers, both for wheat and other arable crops. The majority of farmers sowed wheat at either 25 mm (12% of responses) or 38 mm (69%). This suggests that a recommendation to sow wheat at around 40 mm would be acceptable to most farmers.
The physical and chemical properties of arable soils vary widely, and are likely to affect the practicality of sowing at particular depths, the effect of depth on yield, and the effect of depth on density of exposed seed. This implies a need for flexibility in making recommendations to farmers about sowing depth, to allow them to take account of local conditions.
This variability also implies that there will be some conditions under which seed incorporation is poor, so there is a need for methods to reduce the density of exposed seed after sowing when necessary. We therefore tested the use of rolling and harrowing to reduce seed densities after sowing (Pascual, Saunders, Hart & Mottram, in press). Three experimental field trials were carried out in peaty soil fields in fenland in Eastern England. Both rolling and harrowing buried a large proportion of exposed seeds after sowing or broadcasting, but their relative efficacy was affected by seedbed condition. In a fine seedbed, the proportion of seeds covered by harrowing (84-86%) was substantially higher than that for rolling (53-58%), whereas in a rough seedbed the results were more similar. Harrowing proved very effective for covering simulated spills of seed irrespective of seedbed condition, as it dispersed most spills and buried most of the exposed seeds (85-92%). Rolling was much less effective in dispersing spills. Analysis of data obtained in conjuction with the Pesticide Usage Survey indicates that 77% of farmers already roll after sowing wheat, whereas fewer than 1% harrow (Garthwaite, pers. comm.). This suggests that harrowing large areas is unlikely to gain acceptance by farmers, and that it is more practical to recommend rolling as a standard practice (except when precluded by adverse soil conditions). To disperse small spills of seed, the use of a hand fork or rake would be more practical and effective.
The results of this project show that the measures discussed above would be effective in reducing densities of exposed wheat seed in UK conditions. It is reasonable to assume that this would reduce the rate at which birds could feed on the seed, and therefore the risk of mortality, although we have not confirmed this experimentally. Similar approaches could be considered for other types of seed and other pesticides, although in some cases they will be impractical (e.g. where sowing must be shallow) or the risk will remain high (for example where only one or a few seeds contains a lethal dose). Therefore, decisions on risk management must always take account of the specific ecotoxicological and agronomic factors for the situation concerned.
CONCLUSIONS
This study has identified a range of factors affecting the risk to pigeons feeding on wheat seed treated with fonofos (Table 1). Among the most important are the area sown, the toxicity of the pesticide, its concentration on the seed, the density of exposed seed, the availability of other foods and the ability of birds to selectively avoid treated seed.
The avoidance response was shown to be complex. Its effectiveness in preventing poisoning depends crucially on whether birds stop feeding before ingesting a lethal dose. This depends on feeding rate, which in turn depends on several factors including hunger, daily feeding patterns and group size. Even when a lethal dose is ingested, some birds regurgitate enough to survive.
Taken together these factors imply that fonofos-treated wheat will poison birds but only when both feeding rate and the concentration of residues are high. This is most likely to occur when seed is spilt before sowing, as this produces dense patches of seed which enable rapid feeding, and by-passes losses of pesticide which may be occurring during the sowing process. It could also occur after sowing if large amounts of seed remain uncovered by soil, while the residues remain high. Overall the densities of exposed seeds are such that fewer than 1% of Woodpigeons achieve intake rates over 2 g minute-1, and this must be combined with high residues of fonofos to overcome the avoidance and regurgitation responses and cause mortality. The risk of poisoning is therefore low. This assessment is consistent with evidence from poisoning incidents, roost surveys and radio-tracking, which confirm that the frequency of poisoning is very low.
The central role of feeding rate also helps identify practical methods for farmers to reduce the risks to birds from fonofos-treated wheat. The incorporation of sown seed can be improved by increasing the depth of sowing, and by rolling after sowing. Where spills occur they are best dispersed by harrowing, or the use of a fork or rake if they are small enough.
Some of the results have direct practical application in assessing and managing risks to birds. It has been known for many years that some pesticides are strongly avoided by captive birds, but there has been no generally-accepted method for testing avoidance. The results of this project are currently being used to develop improved methods, which take account of the importance of feeding rate. It is hoped that this will lead to the adoption of an internationally-recognised guideline for avoidance testing. A second practical application has been the use of the results by the UK Pesticides Safety Directorate, to refine their approach to managing risks to birds from seed treatments. In particular, they have adopted new phrases advising farmers to reduce risks by sowing cereal seed at 40 mm and, if excessive seed or spills are present on the soil surface, harrowing and rolling to bury it. These phrases will be placed on the seed bag labels for those pesticides for which regulatory assessment indicates a significant risk.
As stated at the outset, this project focussed on the risk of mortality from short-term exposures of pigeons to fonofos-treated wheat. The results have significantly improved our understanding of these risks, and have direct practical implications for methods of risk assessment and risk management. The principles are likely to apply to short-term exposures for other species and seed treatments, although it must be remembered that the relative importance of different factors will vary (e.g. avoidance will be ineffective for treatments where one or a few seeds carry a lethal dose). Finally, the results illustrate the general importance of understanding the factors which influence risk, including bird behaviour and ecology as well as environmental and agricultural variables.
ACKNOWLEDGEMENTS
We are grateful to Dr A.S. Cooke for advice and information, and for collaborating with us in analysing his observations from Woodwalton Fen. Also to farmers and landowners for co-operation and access to study sites. Zeneca Agrochemicals kindly provided samples of fonofos for use in aviary experiments. The majority of the work was funded by the UK Pesticides Safety Directorate, and we benefited from valuable discussions with a number of their staff, especially Mark Clook. Dr J A Pascual’s contributions and the experiments on manipulation of sowing depth were funded by post-doctoral fellowships from the European Union AIR Programme and the Spanish Government.
REFERENCES
Avery, M.L., Fischer, D.L. & Primus, T.M. 1997. Assessing the hazard to granivorous birds feeding on chemically-treated seeds. Pesticide Science 49: 362-366.
Cooke, A.S. 1988. Poisoning of Woodpigeons on Woodwalton Fen. In: Greaves, M.P., Smith, B.D. & Greig-Smith, P.W. (eds) Field studies for the study of environmental effects of pesticides; Thornton Heath; British Crop Protection Council: 297-301.
Fryday, S.L., Chandler-Morris, S.A. & Hart, A.D.M. 1998. Presentation method and the avoidance of fonofos-treated seed by captive birds. Bulletin of Environmental Contamination and Toxicology 61: 448-454.
Fletcher, M.R., Hunter, K., Barnett, E.A. & Sharp, E.A. 1997. Pesticide poisoning of animals 1996: investigations of suspected incidents in the United Kingdom. Report of the Environmental Panel of the Advisory Committee on Pesticides. London; MAFF: 50pp.
Garthwaite, D.G., Thomas, M.R. & Hart, M. 1995. Pesticide usage survey report 127 – arable farm crops 1994. London; MAFF: 97pp.
Hart, A.D.M. & Thompson, H.M. 1995. Significance of regurgitation in avian toxicity tests. Bulletin of Environmental Contamination and Toxicology 54: 789-796.
Hill, E.F. & Camardese M.B. 1986. Lethal dietary toxicities of environmental contaminants and pesticides to Coturnix. Fish and Wildlife Technical Report 2. Washington DC; US Department of the Interior, Fish and Wildlife Service: 147pp.
Inglis, I.R., Isaacson, A.J., Smith, G.C., Haynes, P.J. & Thearle, R.J.P. 1997. The effect on the Woodpigeon (Columba palumbus) of the introduction of oilseed rape into Britain. Agriculture. Ecosystems & Environment 61: 113-121.
de Leeuw, J., Gorree, M., de Snoo, G.R., Tamis, W.L.M., van der Poll, R.J. & Luttik, R. 1995. Risks of granules and treated seeds to birds on arable fields. Leiden: Leiden University: 131pp.
McKay, H.V., Prosser, P.J., Hart, A.D.M., Langton, S.D., Jones, A., McCoy, C., Chandler-Morris, S.A. & Pascual, J.A. In press. Do pigeons avoid pesticide-treated cereal seed? Journal of Applied Ecology.
Mineau, P., Jobin, B. & Baril, A. 1994. A critique of the avian 5-day dietary test (LC50) as the basis of avian risk assessment. Technical Report No. 215. Hull: Canadian Wildlife Service: 23pp.
Murton, R.K. & Vizoso, M. 1963. Dressed cereal seed as a hazard to wood-pigeons. Annals of Applied Biology 52: 503-517.
Pascual J.A. & Hart, A.D.M. 1997. Exposure of captive Feral Pigeons to fonofos-treated seed in a semi-field experiment. Environmental Toxicology and Chemistry, 16: 2543-2549.
Pascual, J.A., Hart, A.D.M. & Fryday, S.L. In press. Incidence of lethal bird poisoning reduced by the regurgitation of pesticide-treated food. Environmental Toxicology and Chemistry, vol 18, no. 2.
Pascual, J.A., Hart, A.D.M., Saunders, P.J., McKay, H.V., Kilpatrick, J. & Prosser, P. In press. Agricultural methods to reduce the risk to birds from cereal seed treatments. I. Sowing depth manipulation. Agriculture, Ecosystems and Environment.
Pascual, J.A,, Saunders, P.J., Hart, A.D.M. & Mottram, J. In press. Agricultural methods to reduce the risk to birds from cereal seed treatments. II. Rolling and harrowing as post-sowing cultivations. Agriculture, Ecosystems and Environment.
Stanley, P.I. & Bunyan, P.J. 1979. Hazards to wintering geese and other wildlife from the use of dieldrin, chlorfenvinphos and carbophenothion as wheat seed treatments. Proceedings of the Royal Society, London. B 205: 31-45.
Table 1. Relative importance of factors affecting risk, and options for risk management. The effect of each factor is also indicated, e.g. increasing pesticide usage leads to increased risk (shown as ‘+’). The importance of each factor was assessed qualitatively, based on the experimental results and other information reviewed in preceding sections.
Fig. 1. Effect of feeding rate on total seed eaten and risk of mortality, for captive Feral Pigeons feeding on fonofos-treated wheat (Fryday et al., unpublished manuscript). Each point represents the mean of several replicates of one experimental treatment. Percentages show overall mortality in each treatment. Captions within figure indicate the nature of the experimental treatment conditions associated with low and high levels of risk. Mean measured residues on the wheat ranged between 693 and 931 mg.kg-1.
.
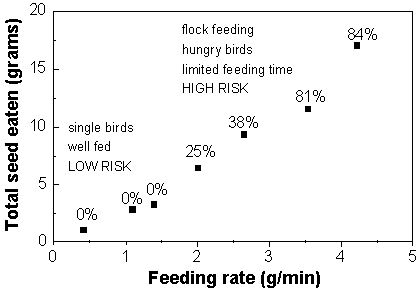
Fig. 2. Conceptual model of the inter-relationships between factors determining the risk of poisoning to birds feeding on a field sown with treated seed. Note that the primary source of risk is from seeds exposed on the soil surface. The contribution of buried seeds to risk is considered negligible, because the time taken to find them increases the opportunity for avoidance to operate (see text).
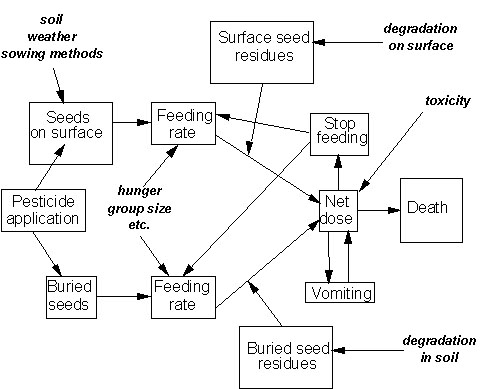
Fig. 3. Illustration of the relationship between feeding rate, pesticide concentration and the risk to birds feeding on treated seed.