S17.5: Ecological barriers and the flexible orientation system of migrating birds
Susanne Åkesson & Roland Sandberg
Department of Animal Ecology, Lund University, Ecology Building, SE-223 62 Lund, Sweden, fax 46 46 2224716, e-mail Susanne.Akesson@zooekol.lu.se; Roland.Sandberg@zooekol.lu.se
Susanne Åkesson & Roland Sandberg. 1999. Ecological barriers and the flexible orientation system of migrating birds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 1005-1016. Johannesburg: BirdLife South Africa.Migratory birds regularly engage in movements which conspicuously deviate from the intended direction of migration. An analysis of ringing recoveries from a coastal site in South Sweden revealed that temporary reverse movements shortly after capture were widespread among several different species of passerine migrants in autumn. Reverse movements occurred more frequently in species with relatively small fat reserves at capture compared to species with large fat deposits. This finding is in agreement with orientation cage experiments with two species of passerine migrants captured at a nearby coastal site in South Sweden. Ringing recoveries of the same and related species showed that the birds did perform temporary reverse movements more frequently at the coastal site as compared to an inland site. Recently, observations of nocturnally migrating passerines recorded by infrared camera and tracking radar along the French and Spanish Mediterranean coasts show important deviations from the intended migratory course associated with the topography of the coast. Increasing proportions of birds performing reverse migration, at low altitudes and at low flight speeds, were observed during the course of the night, suggesting a decreasing preparedness to cross the barrier and possibly indicating a search for suitable stopover sites at dawn. Reverse movements presumably are of adaptive significance for birds confronted with the sea which are predisposed to refuelling. Movements away from the coast will take the bird from an area with high competition for food and high risk of predation to more suitable resting grounds further inland before migration is resumed.
INTRODUCTION
Birds on migration regularly engage in flights deviating from or directed opposite to their intended migratory direction in spring as well as in autumn (e.g. von Haartman 1945; von Haartman et al. 1946; Koskimies 1947; Myres 1964; Evans 1968; Able 1977; Richardson 1978; Lindström & Alerstam 1986; Sandberg 1994; Åkesson et al. 1996; Bruderer 1997; Bruderer & Liechti in press). Orientation errors, effects of wind drift or unsuitable weather has been suggested to explain this behaviour (reviewed by Richardson 1982). In his review, Richardson concluded that different causes might be involved under different environmental circumstances. In areas bordering to ecological barriers (e.g. coasts and deserts) birds that have small fuel reserves when they encounter the barrier on migration might perform temporary reverse movements. The reverse flights might be performed in order to find suitable resting grounds for refuelling before migration is resumed (Alerstam 1978). At the border to the barrier, i.e. at the coast, birds may encounter low food availability as well as experiencing high inter- or intra-specific competition for food (cf. Moore & Yong 1991). Furthermore, the birds may experience a high risk of predation at stopover sites in coastal areas (cf. Lindström 1990). Support for these findings have been found in studies of a number of different species of passerine migrants in South Scandinavia (Lindström & Alerstam 1986; Sandberg et al. 1988; Åkesson et al. 1996; see also Åkesson 1997).
The widespread occurrence of temporary reverse migration among widely different species of passerine migrants is most interesting and suggests an adaptive explanation to reverse movements in migrating birds. However, several interesting questions need to be answered in order to understand the cause of temporary reverse movements of migrants in coastal areas. For instance, is reverse migration a general phenomenon among birds on migration independent of previous experience and regardless of the location of the capture site? Do birds predisposed to refuelling and grounded at inland sites perform reverse migration in order to locate suitable resting grounds, as has been shown for birds in coastal areas? How do these findings correspond with behaviours registered in orientation cages? In this paper we would like to review the most recent findings on avian orientation in relation to geographical barriers and how the birds’ body condition may affect the orientation behaviour when confronted with trans-barrier flights. Many of the studies that have focused on the phenomena of reverse movements and orientation have been performed in South Scandinavia, but more recent reports on the orientation behaviour of free-flying migrants stem from infrared observations and radar tracking in the Mediterranean area (Bruderer 1997; Bruderer & Liechti 1998).
TEMPORARY REVERSE MIGRATION
The occurrence of temporary reverse migration in autumn has been investigated by analysing the direction of migration based on ringing recoveries of passerine migrants in Sweden (Sandberg et al. 1988; Åkesson et al. 1996; Åkesson unpublished manuscript). Extensive ringing data from two of the main Bird Observatories located at a coastal site on the Falsterbo peninsula in southwesternmost Sweden (Falsterbo Bird Observatory: 55°23’N, 12°50’E) and at an inland ringing site at Lake Kvismaren (Kvismare Bird Observatory: 59°10’N, 15°25’E) in south central Sweden were analysed with respect to direction of migration (Åkesson in press). The autumn ringing recoveries within the first calendar year was analysed and divided into two time periods, shortly after capture (Kvismaren: <14 days, Falsterbo: <10 days) and later after capture (Kvismaren: >15 days, Falsterbo: >11 days), respectively. The different lengths of the time periods between the two sites were chosen because at Kvismaren only a limited number of birds was recovered within 10 days after capture.
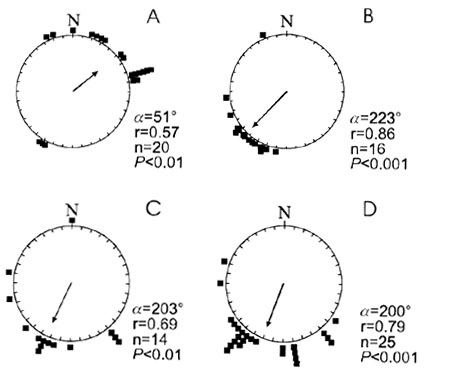
The analysis of ringing recoveries (n = 725) of 20 species of passerines ringed at Falsterbo Bird Observatory revealed that migrating passerines regularly engage in temporary reverse movements in autumn (Åkesson et al. 1996; see also Sandberg et al. 1988). A large proportion of diurnal, as well as nocturnal, short-distance and long-distance migrants perform temporary reverse movements shortly after capture at Falsterbo (64% of all birds recovered <10 days after capture). This finding is examplified for three species of tropical migrants (Whitethroat Sylvia communis, Garden Warbler S. borin and Willow Warbler Phylloscopus trochilus) in Fig. 1A. Recoveries of the same species reported 11 days or later after capture, were found mainly in the direction of natural migration towards south-west at this site (Roos 1984; Fig. 1B). Autumn recoveries reported for a number of tropical migrants ringed at an inland site at Lake Kvismaren in south central Sweden, did not reveal such a pattern of initial temporary reverse migration shortly after capture (Fig. 1C), as was found at the coastal site (Åkesson in press). There was no difference in mean orientation between the two time periods at Kvismaren (Mardia’s one-way classification test: F1,36 = 0.065, P > 0.05; Mardia 1972) (Fig. 1C and Fig 1D). However, the difference in mean orientation for the two groups of tropical migrants recovered shortly after capture at the inland and the coastal site, respectively, was significant (Mardia’s one-way classification test: F1,32 = 38.8, P < 0.001; Mardia 1972), demonstrating that reverse flights occurred at the coastal site, but not at the inland site. These results suggest that temporary reverse flights occur close to barriers and not at inland sites. This behaviour is presumably a response triggered by a confrontation with an ecological barrier such as the sea and is not performed in order to locate known or recently encountered stopover sites when migrants are grounded at inland sites along the migration route. Reverse movements directed opposite to the expected migratory direction, at Falsterbo peninsula coinciding with the direction away from the sea, can be expected to facilitate orientation, since with true reverse orientation the birds do not have to correct for course deviations leading away from an intended migratory route (cf. Alerstam 1990). However, the reverse direction at Falsterbo also coincides with the closest distance to the most suitable resting grounds in this area (cf. Lindström & Alerstam 1986), and therefore based on this data set we cannot conclude that the birds’ direction of movement is selected in order to facilitate orientation.
INFLUENCE OF BODY CONDITION ON ORIENTATION
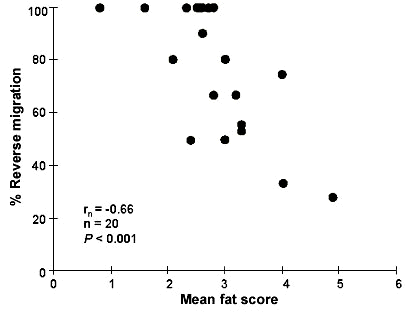
Birds with small fuel deposits that encounter an ecological barrier during migration could be expected either to stop at the barrier in order to refuel or to perform short-distance movements to reach more suitable resting grounds. Because of the high concentration of migrating birds at coastal sites (i.e. expected high competition for food and high risk of predation) the second alternative is probably more plausible. An analysis of ringing recoveries at Falsterbo Bird Observatory revealed that autumn migrating passerines engage in reverse flights and that species carrying relatively small fat deposits at capture (estimated according to a visual classification scale; Pettersson & Hasselquist 1985; fat scores given in Åkesson et al. 1996) more frequently perform reverse migration than do species with large fat reserves (Fig. 2). For two species, the Blue Tit Parus caeruleus and the Reed Warbler Acrocephalus scirpaceus, a relatively high number of recoveries was reported shortly after capture at Falsterbo, enabling a comparison of body mass and fat score of individuals performing reverse migration compared to forward migrating birds. Birds that were recovered in reverse directions carried relatively smaller fat reserves (Reed Warbler) or were lighter (Blue Tits) than individuals reported in forward directions (Åkesson et al. 1996). These findings agree with what has been found for passerines in cage experiments (Sandberg 1994; Bäckman et al. 1997).
The effect of body condition on magnetic directional choices has recently been studied by orientation cage experiments for a nocturnal (Robin Erithacus rubecula, Sandberg 1994) and a diurnal migrant (Chaffinch Fringilla coelebs, Bäckman et al. 1997) in South Sweden. The two species were captured and tested at a coastal site on the island of Öland in the Baltic Sea in south-east Sweden (Ottenby Bird Observatory: 56°12’N, 16°24’E) during autumn migration. For both species a similar behavioural pattern was observed. In the control condition, an experiment performed under clear sky conditions in the local geomagnetic field, the birds showed a bimodal orientation along a north-east/south-west axis (Sandberg 1994; Bäckman et al. 1997). The bimodal orientation closely followed an experimental shift of the magnetic field direction (magnetic north shifted 90° both clockwise and counterclockwise for Robins and 90° counterclockwise for Chaffinches) to north-west/south-east. A subdivision of the data set into two groups based on the body condition (visually estimated size of subcutaneous fat stores), showed that in both the control and experimental group the fat birds oriented in the expected migratory direction towards magnetic south-west. On the other hand, the majority of the lean birds oriented opposite (magnetic north to north-east) to the species’ natural migratory direction at this site. The results clearly show that the birds’ body condition, presumably correlated with the preparedness of forward migration, affects the compass orientation of birds in cages. Similar findings have also been reported for Red-eyed Vireos, Vireo olivaceus, in North America (Sandberg & Moore 1996) indicating a widespread occurrence of reverse migration related to body condition in migrating birds.
THE ORIENTATION BEHAVIOUR OF BIRDS CAPTURED AT FALSTERBO
At Falsterbo, orientation cage experiments, with the same equipment and with the same test procedure as was used for Robins and Chaffinches studied at Ottenby (Sandberg et al. 1988; Sandberg 1994; Bäckman et al. 1997), have not resulted in bimodal orientation that could be explained by the birds’ body condition (e.g. Sandberg et al. 1988; Åkesson 1994; Åkesson & Sandberg 1994). Instead, the birds’ orientation under clear skies more or less coincided with a compromise direction towards the position of the setting sun at test and the expected migratory direction in spring as well as in autumn for a number of passerine migrants (Åkesson 1994; Åkesson & Sandberg 1994), except for one species, the Marsh Warbler Acrocephalus palustris (Åkesson 1993a). The Pied Flycatcher Ficedula hypoleuca is the only species so far that has shown clear responses to experimentally shifted magnetic fields at this site during autumn migration (Åkesson 1994). However, when directional information from the geomagnetic field was cancelled, by exposing the birds to a vertical magnetic field during experiments under overcast and clear sky conditions, the orientation of most of the species became random (Åkesson 1994 and references given therein). Thus, the lack of directional information from the geomagnetic field obviously caused disorientation even though the birds in one experimental condition had access to visual sunset cues for orientation.
Within the different species the orientation behaviour of individual birds cannot be explained by the birds’ energetic status. The Marsh Warbler, which deviates in preferred migratory orientation compared to the other birds tested at Falsterbo, migrates toward south-east in autumn and put on large fat reserves before departing on migration from South Sweden. Contrary to other migrants for which orientation has been investigated at Falsterbo, the orientation of Marsh Warblers does not coincide with the direction towards the sun at sunset (Åkesson 1993a, 1994). Rather, the Marsh Warblers were oriented towards the expected direction of migration towards south-east. Both Marsh and Reed Warblers stopover in reedbeds and wetland areas, which could be found at one of the ringing sites at the Falsterbo peninsula (Roos 1984; Karlsson 1993). Compared to other migrants at Falsterbo they both put on large fat reserves before autumn migration is initiated. Furthermore an analysis of ringing recoveries from Falsterbo showed that Reed Warblers rarely engage in temporary reverse flights, but are mainly recovered in the forward direction towards south-west in autumn (Åkesson et al. 1996), similar to what was found for Marsh Warblers in cage experiments (Åkesson 1993a). For the large part of nocturnal passerine migrants arriving at Falsterbo in autumn suitable resting grounds are limited, and it is presumably adaptive to move away from the coast to inland stopover sites (cf. Alerstam 1978; Lindström & Alerstam 1986; Åkesson et al. 1996). These findings are interesting and suggest that bird species that are able to find suitable resting grounds at the coast do not show sunset attraction in orientation cages (Åkesson 1993a) and mainly depart from the coast in the expected migratory direction (Åkesson et al. 1996). Other migrants, presumably recently confronted with the sea before capture, carry smaller fat reserves when captured and to a high degree perform reverse flights (Åkesson et al. 1996; Fig. 2). In cage experiments they are also affected by the position of the setting sun (cf. Åkesson 1994; Åkesson & Sandberg 1994), which might be related to reduced motivation to continue migration in the expected migratory direction.
OBSERVATIONS OF FREE-FLYING MIGRANTS
Birds on migration are thought to rely on an endogenous program, based on a time sense and an inherited compass course, to find their population-specific breeding areas or wintering grounds (for review see Berthold 1996). Unexpected deflections from the preferred course because of local topography or wind might result in disturbances of such a migration program. Temporary reverse migration and flight directions deviating from the seasonally appropriate migratory directions are well-known phenomena in widely different areas of the globe (for reviews see e.g. Richardson 1982; Alerstam 1996; Bruderer 1997), which might have serious implications on the birds’ orientation system and course control during migration. Fortunately, recent methodological developments (satellite telemetry) permit recording of migration routes selected by large-sized individual birds. Such recordings are important in order to understand the evolution of migration routes and the integration of overall compass courses with local topography in the birds’ orientation system (Alerstam 1996). Several examples of migration routes by large-sized birds have been presented in the literature, which can be evaluated in relation to the basis on which the birds selected their compass courses and how topography might have influenced their orientation (for an extensive review see Alerstam 1996). Recordings of detailed flight trajectories by surveillance radar of spring migrating Knots, Calidris canutus, in South Scandinavia show that the birds migrated on a broad front over southern Scandinavia towards their high arctic breeding grounds and changed their courses counterclockwise during the maximum 400 km route over which they were followed (Gudmundsson 1994). The average flight direction, towards east of north-east (63°), did not follow a rhumbline or great circle route from the wintering areas to the high arctic breeding grounds, as was expected based on alternative compass systems. Neither did the migratory direction of Knots seem to be affected by wind or weather systems (Gudmundsson 1994). Instead, the orientation of these waders seemed to be, on a large-scale, affected by the overall topography of this region whereas minor topographical factors like coastlines and islands did not seem to affect their flight course. Furthermore, long-distance migration routes have been recorded, for example, for migrating Whooping Cranes Grus americana by aerial radiotracking (Kuyt 1992) and White Storks Ciconia ciconia and Brent Geese Branta bernicla hrota, by satellite telemetry (Berthold et al. 1992, 1995; Gudmundsson et al. 1995; Berthold & Querner 1998), revealing that landscape features are clearly involved in the evolution of flight route selection in these species. For all these examples of flock-migrating birds, the navigation system seems to contain short-distance responses to local landscape features as well as long-distance guidance in relation to large-scale topography (Alerstam 1996). Do we find similar effects of topography within passerine migration systems?
For practical reasons, we know much less about how individual small passerines on migration are integrating topography vs. long-distance compass course control for orientation during migration. However, recent ceilometer observations, cage experiments and analyses of ringing recoveries in South Scandinavia (Åkesson 1993b, in press; Sandberg 1994; Åkesson et al. 1996; Bäckman et al. 1997; cf. above) as well as observations of nocturnal passerine migrants by moon-watching and tracking radar along the Alps in south central Europe (Liechti & Bruderer 1986; Liechti et al. 1996 a, b), infrared camera and tracking radar observations in the coastal Mediterranean area (reviewed by Bruderer 1997; Bruderer & Liechti 1998) have produced new information concerning the influence of topography and body condition on passerine orientation. Bruderer & Jenni (1990) and Bruderer (1997) have reviewed the influence of mountain ridges on passerine nocturnal migration in the Swiss Alps. The effect of mountain topography on the birds’ orientation seems to be increasing with an increasing headwind component, cloud cover and decreasing flight altitude (Bruderer 1996). Birds that were captured on Alpine passes mainly stem from northern European populations and those birds carried larger fat reserves than conspecifics captured in lowland areas (Bruderer & Jenni 1990), thus suggesting an increased preparedness to cross the barrier for migrants caught in active flight on Alpine passes (cf. Dorka 1966). However, similar influences of the sun on orientation of passerines, as have been recorded in cage experiments at Falsterbo (Sandberg et al. 1988; Åkesson 1994; Åkesson & Sandberg 1994), have been reported for Chaffinches captured during active migration flights on an alpine pass in South Switzerland (Muheim et al. 1998). Whether this orientation behaviour relates to low motivation to continue migration in the expected migratory direction, or is caused by the experimental method applied (cf. Wiltschko et al. 1997) remains to be investigated. It would be interesting to know if temporary reverse migration occurs in areas bordering mountain barriers, as has been reported for migrants in coastal areas (Sandberg et al. 1988; Åkesson et al. 1996; Bruderer & Liechti 1998).
Earlier studies of passerine nocturnal migration in coastal areas by surveillance radar (reviewed by Lack 1962; Eastwood 1967), reported minor influence of the outline of the coast on the birds’ flight paths when flying across the coast on migration. Only under particular weather conditions did the birds fly along the coastline. This might partly be caused by the amount by which the orientation of the coast is deviating from the intended flight route (for theoretical considerations of when birds should fly along coasts, see Alerstam & Pettersson 1977). Several other studies have reported that nocturnal migrants follow the overall topography of the coast (Lowery & Newman 1966) and that, in particular, birds fly along coasts when they are at risk of being drifted off-shore by winds (e.g. Drury & Nisbet 1964; Nisbet & Drury 1967; Williams et al. 1972; Alerstam & Pettersson 1977; Richardson 1978; see also Bingman et al. 1983). Directional flight responses to the local topography of coastlines at night have also been reported for autumn migrating passerines flying at low altitudes (approximately <100 m) at two nearby observation sites, on the Falsterbo peninsula in South Sweden (Åkesson 1993b). At one of these observation sites reverse migration was observed during the dark hours prior to sunrise (Åkesson 1995; see also Åkesson 1997), demonstrating that nocturnal migrants initiate flights opposite to the direction of migration and that they presumably are less prone to cross the sea barrier during the second half of the night. At least partly a reduced motivation to cross the sea at the end of the night might be caused by limited fat reserves, since at this time the birds probably are arriving at the coast after several hours of flight. Another reason might be avoidance of predation risk out over the sea, for example, by gulls or birds of prey during daytime.
An increased proportion of birds performing reverse flights in coastal areas during the course of the night has also been reported for nocturnal passerine migrants in the south-western Mediterranean area, at Mallorca and at the Spanish coast near Malaga (reviewed in Bruderer 1997; Bruderer & Liechti 1998). The birds which were engaged in reverse flights flew at lower levels and with lower flight speeds compared to forward migrating birds (Bruderer & Liechti 1998), suggesting the possibility that the descending birds were initiating a search for stopover sites a bit inland from the coast. Contrary to these findings Myres (1964) reported on dawn ascent behaviour and reorientation in nocturnal passerine migrants over the North Sea and north-east Atlantic Ocean in autumn. Thus, the ascending and descending flights might be related to the birds’ passage over sea and land, respectively. By placing the tracking radar close to the coast Bruderer, Liechti and co-workers were able to track the birds’ immediate reaction to the coastline when arriving at the coast after flights over land (for location of observation sites see Bruderer & Liechti 1998). During the first part of the night they observed no immediate reactions to the coastline. However, at one of the observation sites, near Malaga, an increasing proportion of the birds shifted their course from the main migratory direction in this area towards south of south-west to south-west to coincide with the east-west orientation of the coastline (Bruderer & Liechti 1998). This shift in orientation occurred simultaneously over water and land during the second part of the night. The proportion of reverse flights also increased at both sites with the course of the night, further indicating a reduced motivation to cross a barrier of unknown dimensions in daylight and presumably with low fuel reserves (Bruderer & Liechti 1998).
CONCLUDING REMARKS
Based on a number of recent reports reviewed in this paper, the influence of topography, i.e. orientation of coastlines and mountain ridges, on the directional choices of migrating birds must be clear. The body condition of the birds seems to be of great importance for selection of migratory directions in areas bordering ecological barriers (Sandberg 1994; Åkesson et al. 1996; Sandberg & Moore 1996; Bäckman et al. 1997). Although the influence of topography on bird orientation is convincing, several interesting questions related to individual performances still remain to be investigated. For example, what principles of navigation does a migrating bird use during flight? How does the navigation system of birds cope with large unexpected detours from an intended course? We must confess that, despite a large amount of data accumulated, describing migratory routes for a large number of species in widely different geographical areas, we know very little about the mechanisms by which birds orient during migratory flights and by which means the birds correct for detours caused, for example, by wind drift or local topography (cf. displacement experiment by Perdeck 1958). In theory, unexpected displacements might lead to difficulties for the birds to keep an intended migration route (e.g. Alerstam 1990; Richardson 1990). To be able to correct for detours, birds must possess a sense of position or rely on ego-centric navigational principles (cf. Wehner 1998). An interesting possibility is that birds might use information from the geomagnetic field for geographical positioning (i.e. gradient map based on total field intensity and angle of inclination; Lohmann & Lohmann 1996; cf. Wallraff 1990; Wiltschko & Wiltschko 1995; Phillips 1996), as has recently been suggested for Silvereyes Zosterops l. lateralis (Munro et al. 1997) and Homing Pigeons Columbia livia (Weindler et al. 1998). Such a bi-coordinate geomagnetic navigation mechanism would enable birds to correct for unintended detours and locate migration goals along the route as well as at the final destination (cf. the moving goal area concept by Rabøl 1978, 1985). However, one open question still remains: on what scale such a geomagnetic position mechanism might be useful and in which areas of the globe it might work? A recent analysis of flight routes recorded by satellite transmitters for foraging Wandering Albatrosses, Diomedea exulans, in the South Atlantic Ocean (Jouventin & Weimerskirch 1990; Prince et al. 1992; Weimerskirch et al. 1993), revealed that a bi-coordinate navigation mechanism based on two parameters of the geomagnetic field might in theory be possible in some areas of the Atlantic Ocean, but not in other parts (Åkesson & Alerstam 1998). However, until we know the function and sensitivity of a magnetoreception mechanism in birds we cannot exclude a global use for geographic positioning based on geomagnetic information, although in some areas such a system seems unlikely.
ACKNOWLEDGMENTS
This study was financed by grants from the Swedish Natural Science Research Council (NFR) to S. Åkesson and from NFR and Swedish Foundation for International Co-operation in Research and Higher Education (STINT) to R. Sandberg.
REFERENCES
Able, K.P. 1977. The orientation of passerine nocturnal migrants following offshore drift. Auk 94: 320-330.
Åkesson, S. 1993a. Effect of geomagnetic field on orientation of the Marsh Warbler, Acrocephalus palustris, in Sweden and Keyna. Animal Behaviour 46: 1157-1167.
Åkesson, S. 1993b. Coastal migration and wind drift compensation in nocturnal passerine migrants. Ornis Scandinavica 24: 87-94.
Åkesson, S. 1994. Comparative orientation experiments with different species of passerine long-distance migrants: effect of magnetic field manipulation. Animal Behaviour 48: 1379-1393.
Åkesson, S. 1995. Avian migratory orientation: geographic, temporal and geomagnetic effects. PhD Thesis, Lund University, Alnarp, Sweden.
Åkesson, S. 1997. Bimodal orientation and the adaptive significance of temporary reverse bird migration in autumn. In: RIN 1997: Orientation & Navigation - Birds, Humans and Other Animals. Oxford; The Royal Institute of Navigation: 28-1 - 28-9.
Åkesson, S. & Alerstam, T.. 1998. Oceanic navigation: are there any feasible geomagnetic bi-coordinate combinations for albatrosses? Journal of Avian Biology 29: 618-625.
Åkesson, S. & Sandberg, R. 1994. Migratory orientation of passerines at dusk, night and dawn. Ethology 98: 177-191.
Åkesson, S., Karlsson, L., Walinder, G. & Alerstam, T. 1996. Bimodal orientation and the occurrence of temporary reverse bird migration during autumn in south Scandinavia. Behavioural Ecology and Sociobiology 38: 293-302.
Alerstam, T. 1978. Reoriented bird migration in coastal areas: dispersal to suitable resting grounds? Oikos 30: 405-408.
Alerstam, T. 1990. Ecological causes and consequences of bird orientation. Experientia 46: 405-415.
Alerstam, T. 1996. The geographical scale factor in orientation of migrating birds. The Journal of Experimental Biology 199: 9-19.
Alerstam, T. & Pettersson, S.-G. 1977. Why do migrating birds fly along coastlines? Journal of Theoretical Biology 65: 699-712.
Bäckman, J., Pettersson, J. & Sandberg, R. 1997. The influence of fat stores on magnetic orientation in day-migrating Chaffinch, Fringilla coelebs. Ethology 103: 247-256.
Batschelet, E. 1981. Circular statistics in biology. New York; Academic Press: 371pp.
Berthold, P. 1996. Control of bird migration. London; Chapman & Hall: 355pp.
Berthold, P., & Querner, U. 1998. Satellite tracking: A new dimension in bird research. Proceedings of Neurobiological Conference in Göttingen 26: 168.
Berthold, P., Nowak, E. & Querner, U. 1992. Satelliten-Telemetrie beim Weissstorch Ciconia ciconia auf dem Wegzug – eine Pilotstudie. Journal für Ornithologie 133: 155-163.
Berthold, P., Nowak, E. & Querner, U. 1995. Satelliten-Telemetrie eines Zugvogels von Mitteleuropa bis in das südafrikanische Winterquartier: eine Fallstudie am Weissstorch Ciconia ciconia. Journal für Ornithologie 136: 73-76.
Bingman, V.P., Able, K.P. & Kerlinger, P. 1982. Wind drift, compensation, and the use of landmarks by nocturnal bird migrants. Animal Behaviour 30: 49-53.
Bruderer, B. 1996. Vogelzugforschung im Bereich der Alpen 1980-1995. Der Ornitologische Beobachter 93: 119-130.
Bruderer, B. 1997. Migratory directions of birds under the influence of wind and topography. In: RIN 1997: Orientation & Navigation - Birds, Humans and Other Animals. Oxford; The Royal Institute of Navigation: 27-1 - 27-10.
Bruderer, B. & Jenni, L. 1990. Migration across the Alps. In: Gwinner, E. (ed) Bird migration physiology and ecophysiology. Berlin, Heidelberg, New York; Springer: 60-77.
Bruderer, B. & Liechti, F. 1998. Flight behaviour of nocturnally migrating birds in coastal areas – crossing or coasting. Journal of Avian Biology 29: 499-507.
Dorka, V. 1966. Das jahres- und tageszeitliche Zugmuster von Kurz- und Langstreckenziehern nach Beobachtungen auf den Alpenpässen Cou/ Bretolet (Wallis). Der Ornitologische Beobachter 63: 165-223.
Drury, W.H. & Nisbet, I.C.T. 1964. Radar studies of orientation of song bird migrants in southeastern New England. Bird-Banding 35: 69-119.
Eastwood, E. 1967. Radar ornithology. London; Metheuen: 278pp.
Evans, P.R. 1968. Reorientation of passerine night migrants after displacement by the wind. British Birds 61: 281-303.
Gudmundsson, G. A. 1994. Spring migration of the Knot Calidris c. canutus over southern Scandinavia as recorded by radar. Journal of Avian Biology 25: 15-26.
Gudmundsson, G.A., Alerstam, T., Benvenuti, S., Papi, F., Lilliendahl, K. & Åkesson, S. 1995. Examining the limits of flight and orientation performance: satellite tracking of Brent Geese migrating across the Greenland ice-cap. Proceedings of the Royal Society London Series B 261: 73-79.
Haartman, L. von 1945. Umschlagende Zugrichtung beim Buchfinken Fringilla c. coelebs L., im Herbst. Ornis Fennica 22: 10-16.
Haartman, L. von, Bergman, G. & Koskimies, J. 1946. Beobachtungen über umschlagende Zugrichtungen der Bachstelze, Motacilla a. alba L., im Herbst. Ornis Fennica 23: 50-62.
Jouventin, P. & Weimerskirch, H. 1990. Satellite tracking of Wandering Albatrosses. Nature, London 343: 746-748.
Karlsson, L. (ed) 1993. Birds at Falsterbo. Report no 150 from Falsterbo Bird Observatory. Anser, Supplement 33. Lund; Grahns: 155pp.
Koskimies, J. 1947. Über den Herbstzug der Bachstelze Motacilla alba in der Gegend von Helsinki. Ornis Fennica 24: 61-79.
Kuyt, E. 1992. Aerial radio-tracking of Whooping Cranes migrating between Wood Buffalo National Park and Arkansas National Wildlife Refuge, 1981-1984. Canadian Wildlife Service, occasional papers no. 74: 1-53.
Lack, D. 1962. Radar evidence on migratory orientation. British Birds 55: 139-158.
Liechti, F. & Bruderer, B. 1986. Einfluss der lokalen Topographie auf nächtlich ziehende Vögel nach Radarstudien am Alpenrand. Der Ornitologische Beobachter 83: 35-66.
Liechti, F., Peter, D., Lardelli, R. & Bruderer, B. 1996a. Die Alpen, ein Hindernis im nächtlichen Breitfrontzug – eine grossräumige Übersicht nach Mondbeobachtungen. Journal für Ornithologie 137: 337-356.
Liechti, F., Peter, D., Lardelli, R. & Bruderer, B. 1996b. Herbstlicher Vogelzug im Alpenraum nach Mondbeobachtungen – Topographie und Wind beeinflussen den Zugverlauf. Der Ornitologische Beobachter 93: 131-152.
Lindström, Å. 1990. The role of predation risk in stopover habitat selection in migrating Bramblings, Fringilla montifringilla. Behavioural Ecolology 1: 24-35.
Lindström, Å. & Alerstam, T. 1986. The adaptive significance of reoriented migration of Chaffinches Fringilla coelebs and Bramblings F. montifringilla during autumn in southern Sweden. Behavioural Ecology and Sociobiology 19: 417-424.
Lohmann, K.J. & Lohmann, C.M.F. 1996. Detection of magnetic field intensity by sea turtles. Nature, London 380: 59-61.
Lowery, G.H. & Newman, R.J. 1966. A continentwide view of bird migration on four nights in October. Auk 83: 547-486.
Mardia, K.V. 1972. Statistics of directional data. London; Academic Press: 357pp.
Moore, F.R. & Yong, W. 1991. Evidence of food-based competition among passerine migrants during stopover. Behavioural Ecology and Sociobiology 28: 85-90.
Muheim, R., Jenni, L. & Weindler, P. 1998. The orientation behaviour of Chaffinches, Fringilla coelebs, caught during active migratory flight in relation to the sun. Ethology.
Munro, U., Munro, J.A., Phillips, J.B., Wiltschko, R. & Wiltschko, W. 1997. Evidence for a magnetite-based navigational "map" in birds. Naturwissenschaften 84: 26-28.
Myres, M. T. 1964. Dawn ascent and re-orientation of Scandinavian thrushes (Turdus spp.) migrating at night over the northeastern Atlantic Ocean in autumn. Ibis 106: 7-51.
Nisbet, I.C.T. & Drury, W.H. 1967. Orientation of spring migrants studied by radar. Bird-Banding 38: 173-186.
Perdeck, A.C. 1958. Two types of orientation in migrating Starlings Sturnus vulgaris and Chaffinces Fringilla coelebs, as revealed by displacement experiments. Ardea 46: 1-37.
Pettersson, J. & Hasselquist, D. 1985. Fat deposition and migration capacity of Robins Erithacus rubecula and Goldcrests Regulus regulus at Ottenby, Sweden. Ringing and Migration 6: 66-76.
Phillips, J.B. 1996. Magnetic navigation. Journal of Theoretical Biology 180: 309-319.
Prince, P.A., Wood, A.G., Barton, T. & Coxhall, J.P. 1992. Satellite tracking of Wandering Albatrosses Diomedea exulans in the South Atlantic. Antarctic Science 4: 31-36.
Rabøl, J. 1978. One-direction orientation versus goal area navigation in migratory birds. Oikos 3: 216-223.
Rabøl, J. 1985. The moving goal area and the orientation system of migrant birds. Dansk Ornitologisk Forenings Tidskrift 79: 29-42.
Richardson, W.J. 1978. Reorientation of nocturnal landbird migrants over the Atlantic ocean near Nova Scotia in autumn. Auk 95: 717-732.
Richardson, W.J. 1982. Northeastward reverse migration of birds over Nova Scotia, Canada, in autumn. Behavioural Ecology and Sociobiology 10: 193-206.
Richardson, W.J. 1990. Wind and orientation in migrating birds: a review. Experientia 46: 416-425.
Roos, G. 1984. Migration, wintering and longevity of birds ringed at Falsterbo (1947-1980). Anser [Suppl 13], Lund (English summary): 208pp.
Sandberg, R. 1994. Interaction of body condition and magnetic orientation in autumn migrating Robins, Erithacus rubecula. Animal Behaviour 47: 679-686.
Sandberg, R. & Moore, F.R. 1996. Migratory orientation of Redeyed Vireos, Vireo olivaceus, in relation to energetic condition and ecological context. Behavioural Ecology and Sociobiology 39: 1-10.
Sandberg, R., Pettersson, J. & Alerstam, T. 1988. Why do migrating Robins, Erithacus rubecula, captured at two nearby stop-over sites orient differently? Animal Behaviour 36: 865-876.
Wallraff, H. 1990. Conceptual approaches to avian navigation systems. Experientia 46: 379-388.
Wehner, R. 1998. Navigation in context: grand theories and basic mechanisms. Journal of Avian Biology 29: 370-386.
Weimerskirch, H., Salamolard, M., Sarrazin, F. & Jouventin, P. 1993. Foraging strategy of Wandering Albatrosses through the breeding season: a study using satellite telemetry. Auk 110: 325-342.
Weindler, P., Becker, M., Holtkamp-Rötzler, E. & Wiltschko, W. 1998. Daily changes of the Earth’s magnetic field intensity results in deviations of homing directions in pigeons. Proceedings of Neurobiological Conference in Göttingen 26: 535.
Williams, T.C., Williams, J.M., Teal, J.M. & Kanwisher, J.W. 1972. Tracking radar studies of bird migration. In: Galler, S.R., Schmidt-Koenig, K., Jacobs, G.J. & Belleville, R.E. (eds) Animal orientation and navigation. Washington DC; NASA SP-262.
Wiltschko, R. & Wiltschko, W. 1995. Magnetic orientation in animals. Berlin, Heidelberg; Springer: 297pp.
Wiltschko, R., Wiltschko, W. & Munro, U. 1997. Migratory orientation in birds: the effects and after-effects of exposure to conflicting celestial and magnetic cues. In: RIN 1997: Orientation & Navigation - Birds, Humans and Other Animals. Oxford; The Royal Institute of Navigation: 6-1 - 6-14.
Fig. 1. The circular diagrams show the direction of recoveries for tropical nocturnal migrants captured at Falsterbo (A and B) and Kvismare (C and D) Bird Observatories in South Sweden. (A) and (C) show migratory directions shortly after capture (Falsterbo: <10 days; Kvismaren: <14 days), while (B) and (D) show directions of birds recovered later after capture (Falsterbo: >11 days; Kvismaren: >15 days). The birds captured at Falsterbo are Whitethroat Sylvia communis, Garden Warbler S. borin and Willow Warbler Phylloscopus trochilus. Recoveries from Kvismare Bird Observatory are represented by White Wagtail Motacilla alba, Yellow Wagtail M. flava, Whitethroat, Garden Warbler, Blackcap S. atricapilla, Willow Warbler and Pied Flycatcher Ficedula hypoleuca. Mean angles of orientation (a ) are indicated by the arrow and mean vectors (r) are drawn relative to the radius of the circle (ranging between 0 and 1). Numbers of recovered individuals (n) are shown as filled squares in the circular diagrams. Significance levels (P) are according to the Rayleigh test (Batschelet 1981). Modified after Åkesson et al. (1996), Åkesson (1997) and Åkesson (in press).
Fig. 2. The proportion of reverse migration (%) for passerines captured and ringed at a coastal site at Falsterbo Bird Observatory in autumn and recovered within 10 days after capture in relation to mean fat score at capture. Each data point represents the values for one species. Statistics are given for a Spearman rank-order correlation. Values of mean fat score for each species are given in Åkesson et al. (1996). Modified after Åkesson et al. (1996).