S17.4: Celestial and magnetic cues in experimental conflict
Roswitha Wiltschko & Wolfgang Wiltschko
Fachbereich Biologie, Zoologisches Institut, J.W.Goethe-Universität, Postfach 11 19 32, D-60054 Frankfurt a.M., Germany, fax 49 69 798 24820, e-mail
wiltschko@zoology.uni-frankfurt.d400.de Wiltschko, R. & Wiltschko, W. 1999. Celestial and magnetic cues in experimental conflict. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 988-1004. Johannesburg: BirdLife South Africa.Experiments testing migrating birds under conflicting magnetic and celestial cues will be reviewed. The results vary considerably - in some cases, celestial cues seem to dominate, in others, the magnetic field proved dominant, and in even others, conflict between cues leads to disorientation. In tests under the natural sky with magnetic North deflected, the majority of birds tested only once oriented as indicated by celestial cues, whereas birds repeatedly tested under these conditions without seeing the cues in their natural relationship between tests generally altered their course and followed the deflected magnetic field. The time until birds responded, however, varied between species, with some birds responding immediately, while others responded only with delay. In Silvereyes, Zosterops lateralis, immediate and delayed responses were observed, depending on the direction in which magnetic North was shifted. This points out a certain asymmetry in response; differences between tests in the Northern and Southern Hemisphere suggest the relationship between the direction of shift and that of celestial rotation as a possible reason. Subsequent tests on the effect of conflicting cues on the birds' response to celestial cues showed that the celestial compass mechanisms had been recalibrated according to the experimental magnetic field, restoring the agreement between cues. There does not seem to be a difference between sunset cues and stars in this respect. Data on the effect of conflicting cues on the magnetic compass are controversial.
INTRODUCTION
During migration, birds fly up to several thousand kilometers from their breeding area to a winter home in a distant region of the earth. For young, inexperienced migrants on their first migration, this means that they must reach a still unknown goal, the species-specific wintering area. They are guided by an innate program that indicates direction and distance of the route (for details, see Berthold 1988). Experienced migrants having established a navigational 'map' during previous migrations, may additionally employ mechanisms of true navigation to head toward their goal (see Perdeck 1958, 1983).
In both cases, compass mechanisms allow birds to locate their migratory direction. These mechanisms are based on two types of factors: the geomagnetic field and celestial cues. While the geomagnetic field provides directional information all the time, the nature of celestial cues varies. During daytime, the sun is the available cue. The sun compass of birds, compensating for the sun's movement with the help of the internal clock, has been analysed in detail in experiments with homing pigeons (e.g. Schmidt-Koenig 1958; Keeton 1974; Schmidt-Koenig et al. 1991; R. Wiltschko et al. 1994). Directional training has indicated the existence of the same mechanism in passerine migrants (Hoffmann 1954; von Saint Paul 1954; Able & Dillon 1977). During migration, however, the sun compass does not seem to play the dominant role that is observed in homing (R. Wiltschko 1980; Orth & Wiltschko 1981; Munro & Wiltschko 1993a). Its use might be largely restricted to the stationary phases when migrants stay in the breeding or wintering area. This is probably due to the fact that the sun's arc varies greatly with geographic latitude and would require a continuous readjustment of the compensation mechanisms as the birds cross latitudes during their extended migration flights (see R. Wiltschko 1981; Munro & Wiltschko 1993a).
The stars at night, in contrast, are of great importance for nocturnal migrants (Emlen 1967a). The star compass does not involve the internal clock nor compensation mechanisms; birds use the configurations of stars and the constant spatial relationships between them for locating directions, just as we are able to locate North with the help of the Great Dipper, Ursa major, irrespective of its position (Emlen 1967b). As a consequence, the star compass is not affected by the latitudinal shifts that migrants experience during migration, thus being an efficient orientation mechanism for extended flights across latitudes. Other prominent celestial cues of great importance for the orientation of nocturnal migrants, many of which start their migration flights at the time of sunset, are provided by the view of the setting sun (Moore 1982, 1985), horizon glow and especially the pattern of polarised light that runs through the zenith at an angle of 90° to the setting sun (Able 1982, 1989). Whether these 'sunset cues' represent a set of independent cues indicating westerly directions, or whether they should be considered as a part of the sun compass, is not entirely clear (for discussion, see Moore 1987). Clock-shift experiments (Able & Cherry 1986; Helbig 1991) appear to suggest the latter.
As migrants generally seem to be able to use both the magnetic field and celestial cues for orientation (see Able 1991; W. Wiltschko & Wiltschko 1991a), the interrelationship of the different sets of cues became a question of great interest: How do the birds rate and rank the various cues? - Experimental evidence suggests that two phases of migratory orientation may be distinguished (see W. Wiltschko et al. 1998a): The first includes the pre-migratory period and the onset of migration. Orientation during this first phase was analysed in series of experiments with birds handraised under various conditions controlling their early experience. Celestial rotation, that is, the rotation of the stars around the celestial pole (Emlen 1970; W. Wiltschko et al. 1987; Able & Able 1990) or the rotation of the diurnal pattern of polarised light (Able & Able 1993; Weindler et al. 1998), proved to be the crucial celestial cue for establishing the migration course, in the case of conflict dominating over information from the magnetic field (see Weindler & Liepa 1999). The second phase includes migration itself, after the birds have left their breeding grounds until they reach their winter quarters, and the return trip back to the breeding ground in spring. Here, in contrast to the first phase, rotation is no longer required. Under experimental conditions, birds now orient with stationary stars as only cues (e.g. Emlen 1970; W. Wiltschko et al. 1987; Weindler et al. 1996). Obviously, the birds are now able to interpret the celestial features themselves and derive directions from them without delay.
The present paper focuses on the interrelation between magnetic and celestial cues during this second phase of migratory orientation, addressing the question how the matured orientation system, at a time when it is actively used, rates and ranks the available cues.
SETTING-UP A CONFLICT BETWEEN THE MAGNETIC AND CELESTIAL CUES
The question about the interrelationship between the various types of cues has been addressed in so-called cue-conflict experiments. The test birds of these studies were wild-caught birds, that is, they had grown up in the wild and had developed their migratory orientation under natural conditions. The vast majority had actually migrated, as they were captured either as transmigrants at stop-over sites or, for spring experiments, in their winter quarters. How would such birds respond when suddenly faced with the familiar set of cues giving contradicting information?
Cue-conflicts can be set up in two ways: one can either manipulate celestial or magnetic cues. Both approaches have been followed up. The majority of studies used the latter design, testing the birds under the natural sky with magnetic North deflected. This is mainly for practical reasons: an artificial magnetic field, produced by a coil system of sufficient size, can be made to resemble the geomagnetic field in almost every detail, except that magnetic North is in a different geographic direction. That is, the birds, being exposed to a rather nature-like stimulus, should not be able to realise a difference from the local geomagnetic field.
Manipulating celestial cues is far more difficult. Analysing the role of the stars, researchers profited from the fact that the natural sky could be replaced by a planetarium sky that allows arbitrary changes in position and composition of stars. Sunset cues were altered by blocking the direct view of the sun and reflecting the sun with mirrors or by changing the pattern of polarised light with polarisers. However, all these methods lead to certain unavoidable changes in the appearance of the sky, and it is not always clear how these might affect the behaviour of birds. One need not be overly concerned when birds tested under a planetarium sky are compared with control birds tested in the same planetarium. In the case of sunset cues, however, the differences in the appearence of the sky are no longer negligible. This is especially true for tests involving polarisers, as the pattern of polarisation produced by polarisers differs markedly from the natural one (see Able 1989). The birds tended to align parallel to the e-vector instead of choosing the angle they normally prefer, often becoming bimodal (see Helbig & Wiltschko 1989 for discussion). These responses to artificial polarisation are fundamentally different from those to natural polarisation and appear to reflect some kind of polarotaxis or phototaxis. The respective results cannot be compared with those of the other cue-conflict experiments; therefore they are excluded from the present considerations.
An elegant method of causing conflict between magnetic and celestial cues without altering the natural appearance of the sky is shifting the birds' internal clock. It is restricted to mechanisms involving time-compensation, and has been successfully used with the sun compass and the sunset cues.
ORIENTATION BEHAVIOUR UNDER CONFLICTING CUES
When birds are tested under conflicting cues, irrespective of the method used, several types of responses appear possible, namely: (1) the birds follow celestial cues, ignoring contradicting magnetic information; (2) they rely on magnetic information and ignore contradicting information from the sky; (3) they might assign equal weight to both types of cues and take up a compromise direction, or (4) they are confused by the sudden discrepancy between formerly agreeing cues, which may lead to disorientation.
All four types of responses have been observed. But although the results appear to be rather diverse at the first glimpse, they form a largely consistent picture, from which certain rules controlling the interactions between the magnetic cues and celestial cues during migration may be inferred.
Experiments under the natural sky in deflected magnetic fields
The tests under the natural sky involved a variety of celestial cues. Testing took place at the time of sunset, with the setting sun and/or the polarisation pattern as crucial cues, others only after dark under the starry sky and one series during the day. The magnetic field was deflected clockwise or counterclockwise by 90° or 120°. Corresponding changes in the birds' preferred direction would indicate orientation by magnetic cues, whereas continuing in the same direction as control birds tested in the geomagnetic field would indicate that birds followed cestestial cues.
According to the test procedures, the cue-conflict experiments fall into two categories:
(1) Individual birds were tested only once under conflicting cues. All these tests were performed at sunset and involved eight species of night migrants. The results, listed in Table 1, are truly mixed. Of the 21 experimental samples, only two (10%) from Snow Buntings, Plectophenax hyperboreus, clearly responded to the deflection of magnetic North with a corresponding change in headings, four (19%), one from Wheatears, Oenanthe oenanthe, one from Pied Flycatchers, Ficedula hypoleuca , and two from Red-eyed Vireos, Vireo olivaceus, showed a small shift, apparently following some kind of compromise between the direction indicated by celestial cues and that indicated by the magnetic field, while ten (48%) oriented as indicated by celestial cues. Five samples (24%) were disoriented, became bimodal or showed an uninterpretable response.
(2) Individual birds were tested repeatedlyunder conflicting cues without being exposed to the two sets of cues in their normal relationship between tests. These tests involved various types of celestial cues, the test birds being night migrants like three species of European Warblers of the genus Sylvia , European Robins, Erithacus rubecula , and Dunnocks, Prunella modularis , as well as Silvereyes migrating at dawn and dusk, and Yellow-faced Honeyeaters, Lichenostomus chrysops, that are day migrants. In this case, the results, listed in Table 2, are rather uniform: All 11 samples followed magnetic cues. A difference between the various types of celestial cues - stars, sunset cues and the daytime sky - is not indicated.
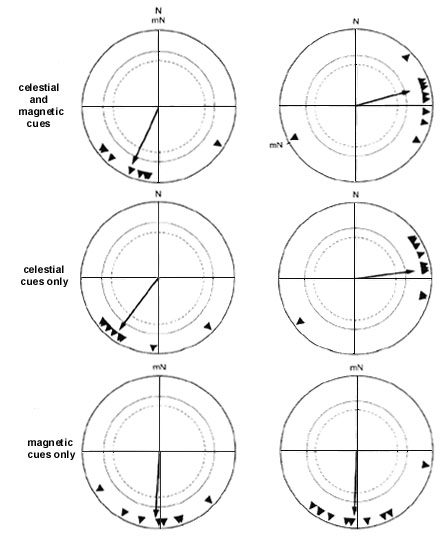
The reasons for this influence of test design on the results become evident upon a closer inspection of the data from the repeated exposures: some birds respond to the deflection of magnetic North only with a certain delay (see Table 2). The European Robin is a typical example. Eight samples of robins tested only once at sunset either preferred their original direction or became disoriented or bimodal (Sandberg et al. 1988a,b; see Table 1). Their behaviour appeared to suggest a dominance of sunset cues over the magnetic field, although the observed bimodality might reflect some kind of confusion. When tested repeatedly at night, robins at first also continued in their original direction, presumably with the help of the starry sky, and seemed to ignore the deflection of magnetic North (Fig. 1, center). In later tests, however, they changed direction and perferred their migratory direction with respect to the experimentally altered magnetic field (Fig. 1, right; W. Wiltschko & Wiltschko 1975b). This implies that the behaviour observed by Sandberg et al. (1988b) might have been a transient stage and that the birds might also have changed their orientation as indicated by the magnetic field if they had been tested more often. This could also apply to some of the other species listed in Table 1 that failed to respond to the change in magnetic North.
A comparison of the studies by W. Wiltschko & Wiltschko (1975b) and Bingman (1987), both showing a delayed response of robins to the deflection of magnetic North, suggests that the number of tests, that is, the number of times the birds were placed into the test cage and thus confronted with conflicting cues, rather than the total duration of the exposure is important. In both studies, robins changed their direction according to magnetic North after two tests. With a recording time of 1 to 1:15 h, this meant a total exposure to conflicting cues of less than 3 h in Bingman's (1987) study, whereas it meant more than 30 h exposure in the study of W. Wiltschko and Wiltschko (1975b), where the birds had been exposed to conflicting cues from 1700 to 1000 next morning.
In contrast to European Robins and Silvereyes, other species listed in Table 2 as well as Snow Buntings and to a certain extent also Pied Flycatchers and Red-eyed Vireos (see Table 1) responded immediately to the altered magnetic field. The reasons for these differences remain unclear. When we first observed the immediate response of European warblers and the delayed response of robins in studies using identical equipment and procedures (W. Wiltschko & Wiltschko 1975a,b), we considered the distance of migration - the warbler species were trans-Saharan migrants, whereas the robins winter in the Mediterranean region -, autumn versus spring migration, and species-specific differences as possible causes. However, a recent study with Silvereyes shows that neither of these possibilities might be the only reason. One group of Silvereyes, tested in a magnetic field deflected 120° counterclockwise to the westsouthwest, responded to the shift in magnetic North immediately with a corresponding change in headings, whereas another group, tested in a field deflected 120° clockwise to eastsoutheast, showed a delayed response, changing their headings only after six to seven exposures to the cue-conflict situation. That is, we observed an immediate and a delayed response in the same species during the same migration season, depending on the direction in which magnetic North was deflected. This indicates a certain asymmetry in response. Akesson (1994), reviewing the cue-conflict experiments under the natural sky reported in the literature, had already called attention to a similar asymmetry: birds appeared to follow a deflection of magnetic North more readily when magnetic North was shifted clockwise than when it was shifted counterclockwise. The data she had considered were all from the Northern Hemisphere; the Silvereye data, obtained in the Southern Hemisphere in Australia indicate a reversed relationship. This suggests that the directional relationship between celestial rotation and the direction in which magnetic North is deflected might also be important for the timing of the birds' response.
In summary, the available data look rather diverse, indicating differences between species. Yet these differences might be only transient; they seem to involve the time until the birds respond to a deflection of magnetic North rather than the rating and ranking of magnetic and celestial cues themselves. Even if it is not clear in all cases, the existing evidence is consistent with the hypothesis that the magnetic field will eventually dominate over celestial cues. When taking up their headings, some birds, however, obviously prefer to use celestial cues; they seem to check these cues against the magnetic field only at intervals and realize any discrepancy between cues with delay, whereas others species appear to pay regular attention to magnetic cues.
Experiments manipulating celestial cues
A considerable number of studies followed the reverse design, testing birds in the local magnetic field while manipulating celestial cues. They include tests under a manipulated planetarium sky, reflecting the setting sun with mirrors and shifting the birds' internal clock at sunset and during the day. Here, a significant change in behaviour indicates that the birds rely on celestial cues, whereas maintaining the same headings as before indicates the use of magnetic cues. The results are listed in Table 3. Again, the outcome appears to be rather mixed. Details of the test design have not always been documented; in particular, the frequency of exposure is often unclear. In general, however, a pattern roughly similar to the one observed when the magnetic field was deflected seems to emerge: Indigo Buntings, Passerina cyanea, Savannah Sparrows, Passerculus sandwichensis, and European Robins tested only once or twice in a cue-conflict situation followed celestial cues or became disoriented, whereas Bobolinks, Dolichonyx oryzivorus, repeatedly tested under a reversed planetarium sky preferred magnetic cues, with some individuals showing delayed responses (Beason 1987). In Moore's (1982) study with Savannah Sparrows, the response is not entirely clear. Despite up to five exposures, the majority of birds seemed to follow sunset cues deflected by mirrors. Whether these findings really indicate a dominance of sunset cues over magnetic cues is unclear, however; in view of the delayed response of Silvereyes after six or seven tests at sunset, they might still reflect a transient stage. Moore (1982) himself pointed out considerable differences between birds, indicating that some individuals might not be using sunset cues.
Clock-shift tests with day migrants did not produce clear evidence of sun compass use. European Starlings, Sturnus vulgaris, during spring migration were the only birds that showed a significant directional deflection, but even here, the shift was smaller than expected for a 6 h shift (W. Wiltschko 1980). The data from Yellow-faced Honeyeaters do not indicate sun compass use. In the first tests, the birds were largely disoriented, but they returned to their original headings from test five or six onward (Munro & Wiltschko 1993a). To a subsequent change in the magnetic field, however, they responded immediately (Munro & Wiltschko 1993b; see Table 2).
Although Homing Pigeons, Columba livia, use their sun compass in a different behavioural context, their response to clock-shifting represents an interesting parallel to the behaviour of migrants in cue-conflict experiments. Pigeons are normally released only once with the sun compass and the magnetic compass giving conflicting information - back in their loft, their photoperiod readjusts to the natural day. In such tests, the vanishing bearings of 6 h clock-shifted pigeons show a characteristic deviation from that of untreated controls mostly in the range of 60° to 120°, depending on time of day and season, which indicates a dominance of the sun compass over the magnetic compass (e.g. Keeton 1974; Neuss & Wallraff 1988; Schmidt-Koenig et al. 1991; R. Wiltschko et al. 1994 a.m.o.). Yet pigeons kept over an extended period in the shifted photoperiod and repeatedly released under these conditions, soon ceased to show the typical deflection and oriented more and more like controls (W. Wiltschko et al. 1984). Like in migrants, the dominance of celestial cues over the magnetic field represents just a transient stage; pigeons repeatedly experiencing conflicting cues eventually gave up their sun compass. - In summary, the two complementary types of cue-conflict experiments produce largely similar results, suggesting that during migration (and homing), the magnetic field is the dominant cue, at least on a long-term basis.
AFTER-EFFECTS OF EXPOSURE TO CONFLICTING CUES
The number of experiments studying possible after-effects of exposure to conflicting cues is rather limited. Such tests are of great interest, however, because the subsequent responses to either set of cues reveal how the birds solved the cue conflict for themselves. There are two obvious possibilities: The birds might simply ignore the one set of cues and rely exclusively on the other, or they might readjust the directional significance of one set of cues to match the other in order to bring the two sets of cues into a new agreement.
After-effects on celestial cues
All studies examining after-effects on celestial cues followed exposures to the natural evening sky or night sky in a deflected magnetic field. When birds were subsequently tested under the natural sky in a vertical magnetic field not providing directional information, the frequency of the previous exposures to conflicting cues again influenced the results. Birds tested only once under conflicting cues at sunset (see Table 1) failed to show a clear effect. Mostly, the ex-experimentals, like the ex-control birds, either preferred the control direction or became disoriented (e.g. Akesson 1994). This is not surprising, since a possible transfer of information between the two cue systems means that the birds first become aware of the cue conflict. Birds must be expected to need time to compare cues, and an information transfer itself might require a certain amount of time.
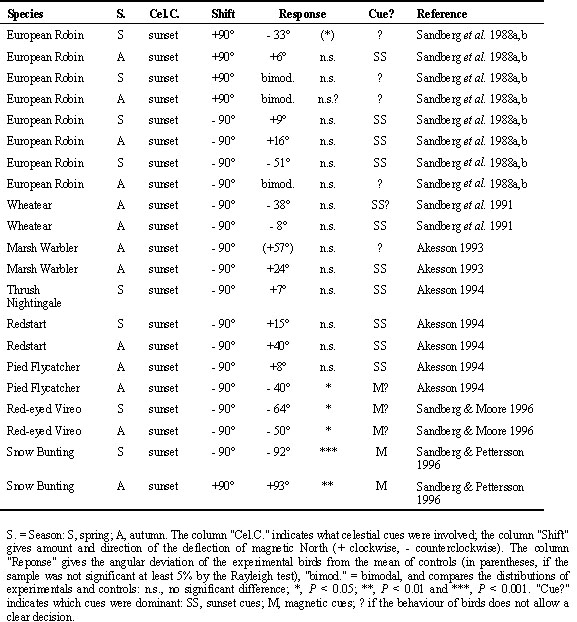
After-effects are thus to be expected only after repeated exposures. The upper section of Table 4 lists the results of the studies looking for possible after-effects on celestial cues by testing birds under the natural sky without magnetic information. During the cue-conflict experiments, the magnetic field had been dominant (see Table 2); when the birds had now to rely on celestial cues alone, they showed the same directional tendencies as previously in the altered magnetic field, that is they continued along their former magnetic course now with the help of celestial cues. Fig. 2 gives, as an example, the respective data from Silvereyes under conflicting cues with magnetic North shifted to 240° westsouthwest (upper diagrams) and from subsequent tests under the natural sky without magnetic information (center diagrams) - the correspondence of the data is striking. This indicates that celestial cues had been recalibrated according to the experimental magnetic field.
The tests with Garden Warblers, Sylvia borin, and European Robins (W. Wiltschko & Wiltschko 1975a,b) involved stellar cues, those with Dunnocks (Bingman & Wiltschko 1988) and Silvereyes (W. Wiltschko et al. 1998b) sunset cues, in particular the pattern of polarised light. Both types of cues seem to be recalibrated in the same way. Interestingly, this seems to be also true for the sun compass of Homing Pigeons. Pigeons kept in a shifted photoperiod for an extended period of time ceased to show the typical deflection from untreated controls; on their first flight after resynchronization with the natural day, they showed a deflection to the opposite direction, indicating that they had recalibrated their sun compass according to the experimental situation (W. Wiltschko
et al. 1984).For recalibration, there was also no difference between birds responding immediately to the change in magnetic North and those responding with delay. Robins needed about two to three exposures to recalibrate their star compass (W. Wiltschko & Wiltschko 1975b). How long the process lasted in Silvereyes is unknown; after at least ten exposures to the cue-conflict situation, when the tests in the vertical magnetic field began, the process had been completed (W. Wiltschko
et al. 1998b).These results clearly show that when faced with a conflict between cues, birds do not merely ignore celestial cues, but recalibrate them to match magnetic information. This might be a general feature of the mature navigation system in migration as well as in homing. Altering the directional significance of celestial cues means that two cue systems are again in harmony, indicating identical directions. Celestial cues and the magnetic field thus appear to be integrated components of a complex mechanism for direction finding (see W. Wiltschko & Wiltschko 1991b).
After-effects on magnetic cuesIn view of these findings, possible after-effects of a cue-conflict on magnetic cues were seldom considered. Only two studies are available so far, one with Savannah Sparrows and one with Silvereyes. The results appear to contradict each other to some degree (see Table 4, lower section). For Silvereyes, the magnetic field had been the dominant cue in the cue-conflict situation (Fig. 2, upper diagrams), and the birds had recalibrated celestial cues (center diagrams), so it is not surprising that their magnetic compass course was unaffected (Fig. 2, lower diagrams; R. Wiltschko et al. 1998). These findings are in accordance with the general picture emerging in long-term cue conflict studies.
Able and Able (1995), who exposed Savannah Sparrows up to five days and nights to the natural sky in a deflected magnetic field, reported a change in the magnetic compass course. This response, indicating a dominance of celestial cues over magnetic cues, contrasts with the other results from cue-conflict studies during migration. Unfortunately, the Ables did not record their birds' directional tendencies during exposure to conflicting cues nor a response to celestial cues alone, so that it is unknown whether their birds' behaviour under those conditions is comparable to that observed in the other studies. The dominance of celestial cues indicated in their study is similar to the relationship between cues normally observed during the pre-migratory period, when innate information on the migratory direction is converted into an actual compass course (see Weindler & Liepa 1999). The reasons are unclear. There are several differences between the study of Able and Able (1995) and the other studies, involving duration and timing of the exposure (see R. Wiltschko et al. 1998 for a detailed discussion). More important, however, might be that the Savannah Sparrows had been caught and were exposed on their breeding grounds, whereas all other long-term studies involved birds caught on migration or near their wintering area. Also, the tests with Savannah Sparrows took place in a magnetic field with a fairly steep inclination, where celestial cues might generally be of increased importance (see Weindler et al. 1995, 1998). Whether the fact that the Savannah Sparrows had not initiated migration under natural conditions or whether the magnetic conditions at the test site are the reason why their response differs from that of the other species or whether species-specific differences are involved, can only be decided by future experiments.
THE ROLE OF MAGNETIC CUES DURING MIGRATION
The findings discussed here indicate a considerable variability between migratory birds concerning the cues on which they base their daily headings, some preferring celestial, others magnetic cues. However, with the possible exception of Savannah Sparrows, even species that prefer to use celestial cues seem to check these cues against the magnetic field at varying intervals and, in the case of cue conflict, orient as indicated by the magnetic field, although often with a certain delay. At the same time, the birds overcome any discrepancy between magnetic and celestial cues by readjusting the directional significance of celestial cues to match the magnetic field, so that both types of cues are again in agreement, providing identical directional information. A difference between the various types of celestial cues in this respect is not indicated.
By generally indicating a dominant role of magnetic cues, the findings on the relationship between magnetic and celestial cues during migration contrast sharply with the results of ontogenetic studies analysing the role of the various cues during the premigratory period, where celestial rotation proved a most important cue, dominating over information from the magnetic field (Emlen 1970; Bingman 1983; Able & Able 1990, 1993; Prinz & Wiltschko 1992). This indicates a fundamental difference between the processes establishing the migratory direction during the pre-migratory period before the onset of migration and orientation en route during migration, which involve two very different tasks. During the pre-migratory period, birds must convert genetically coded information on their migratory direction into an actual compass course. During migration, the respective course is known; it must regularly be located to orient the daily flights. Establishing the migratory course and locating it during actual migration appear to represent two different phases of migratory orientation, apparently controlled by different sets of rules. During both phases, however, a discrepancy between cues is avoided by adjusting one set of cues to the other. Here, celestial and magnetic cues switch their roles, with celestial cues being dominant during the premigratory period and magnetic cues becoming dominant during actual migration.
Possible reasons for this change in dominance between the two cue systems might lie in the nature of cues, in particular in their changes with latitude, as the birds move toward the equator. The dominance of celestial cues appears to cease when the birds no longer depend on celestial rotation, which might become difficult to observe once the birds are en route. As they cover increasing distances, the sky undergoes marked changes due to latitudinal shifts. The familiar stars lose altitude, finally disappearing below the northern horizon, while new stars appear in the south. The magnetic field, in contrast, becomes increasingly regular at lower latitudes (see R. Wiltschko & Wiltschko 1995), providing a reliable source of directional information, well suited to calibrate the newly appearing stars. So it is maybe not surprising that during migration, the birds strongly rely on magnetic information and adjust the directional significance of celestial cues accordingly.
REFERENCES
Able, K.P. 1982. Skylight polarisation patterns at dusk influence migratory orientation in birds. Nature 299: 550-551.
Able, K.P. 1989. Skylight polarisation patterns and the orientation of migratory birds. Journal of Experimental Biology 141: 241-256.
Able, K.P. 1991. Common themes and variations in animal orientation systems. American Zoologist 31: 157-167.
Able, K.P. & Able, M.A. 1990. Calibration of the magnetic compass of a migratory bird by celestial rotation. Nature 347: 378-389.
Able, K.P. & Able, M.A. 1993. Daytime calibration of magnetic orientation in a migratory bird requires a view of skylight polarisation. Nature 364: 523-525.
Able, K.P. & Able, M.A. 1995. Interaction in the flexible orientation system of a migratory bird. Nature 375: 230-232.
Able, K.P. & Cherry, J.D. 1986. Mechanisms of dusk orientation in White-throated Sparrows (Zonotrichia albicollis): Clock-shift experiments. Journal of Comparative Physiology 159: 107-113.
Able, K.P. & Dillon, P.M. 1977. Sun compass orientation in a nocturnal migrant, the White-throated Sparrow. Condor 79: 393-395.
Akesson, S. 1993. Effect of geomagnetic field on orientation of the Marsh Warbler, Acrocephalus palustris, in Sweden and Kenya. Animal Behaviour 46: 1157-1167.
Akesson, S. 1994. Comparative orientation experiments with different species of passerine long-distance migrants: effect of magnetic field manipulations. Animal Behaviour 48: 1379-1393.
Beason, R.C. 1987. Interaction of visual and non-visual cues during migratory orientation by the Bobolink (Dolichonyx oryzivorus). Journal für Ornithologie 128: 317-324.
Berthold, P. 1988. The control of migration in European warblers. In: Ouellet, H. (ed.) Acta XIX Congressus Internationalis Ornithologici. Ottawa; University of Ottawa Press: 215-294.
Bingman, V.P. 1983. Magnetic field orientation of migratory Savannah Sparrows with different first summer experience. Behaviour 87: 43-53.
Bingman, V.P. 1987. Earth's magnetism and the nocturnal orientation of migratory European Robins. Auk 104: 523-525.
Bingman, V.P. & Wiltschko, W. 1988. Orientation of Dunnocks (Prunella modularis) at sunset. Ethology 77: 1-9.
Emlen, S.T. 1967a. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part I: Evidence for use of celestial cues. Auk 84: 309-342.
Emlen, S.T. 1967b. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part II: Mechanisms of celestial orientation. Auk 84: 463-489.
Emlen, S.T. 1970. Celestial rotation: its importance in the development of migratory orientation. Science 170: 1198-1201.
Helbig, A.J. 1991. Dusk orientation of the European Robin: the role of sun-related directional information. Animal Behaviour 41: 313-322.
Helbig, A.J. & Wiltschko, W. 1989. The skylight polarisation patterns at dusk affect the orientation behavior of Blackcaps, Sylvia atricapilla. Naturwissenschaften 76: 227-229.
Hoffmann, K. 1954. Versuche zu der im Richtungsfinden der Vögel enthaltenen Zeitschätzung. Zeitschrift für Tierpsychologie 11: 453-475.
Keeton, W.T. 1974. The orientational and navigational basis of homing in birds. Advanced Study in Behavior 5: 47-132.
Moore, F.R. 1982. Sunset and the orientation of a nocturnal bird migrant: a mirror experiment. Behavioral Ecology and Sociobiology 10: 153-155.
Moore, F.R. 1985. Integration of environmental stimuli in the migratory orientation of the Savannah Sparrow, Passerculus sandwichensis. Animal Behaviour 33: 657-663.
Moore, F.R. 1987. Sunset and the orientation behaviour of migrating birds. Biological Review 62: 65-86.
Munro, U. & Wiltschko, R. 1993a. Clock-shift experiments with migratory Yellow-faced Honeyeaters, Lichenostomus chrysops (Meliphagidae), an Autralian day migrating bird. Journal of Experimental Biology 181: 233-244.
Munro, U. & Wiltschko, W. 1993b. Magnetic compass orientation in the Yellow-faced Honeyeater, Lichenostomus chrysops (Meliphagidae), a day-migrating bird from Australia. Behavioral Ecology and Sociobiology 32: 141-145.
Neuss, M. & Wallraff, H.G. 1988. Orientation of displaced Homing Pigeons with shifted circadian clocks: predictions vs. observation. Naturwissenschaften 75: 363-365.
Orth, G. & Wiltschko, W. 1981. Die Orientierung von Wiesenpiepern (Anthus pratensis L.). Verhandlungen der Deutschen Zoologischen Gesellschaft 1981: 252.
Perdeck, A.C. 1958. Two types of orientation in migrating Sturnus vulgaris as revealed by displacement experiments. Ardea 46: 1-37.
Perdeck, A.C. 1983. An experiment of the orientation of juvenile starlings during spring migration: an addendum. Ardea 71: 255.
Prinz, K. & Wiltschko, W. 1992. Migratory orientation of Pied Flycatchers: interaction of stellar and magnetic information during ontogeny. Animal Behaviour 44: 539-545.
Sandberg, R. 1991. Sunset orientation of Robins, Erithacus rubecula, with different fields of sky vision. Behavioral Ecology and Sociobiology 28: 77-83.
Sandberg, R. & Moore, F.R. 1996. Migratory orientation of Red-eyed Vireos, Vireo olivaceus, in relation to energetic condition and ecological context. Behavioral Ecology and Sociobiology 39: 1-10.
Sandberg, R. & Pettersson, J. 1996. Magnetic orientation of Snow Buntings (Plectrophenax nivalis), a species breeding in the high arctic: passage migration through temperate-zone areas. Journal of Experimental Biology 199: 1899-1905.
Sandberg, R., Pettersson, J. & Alerstam, T. 1988a. Why do migrating Robins, Erithacus rubecula, captured at two nearby stop-over sites orient differently? Animal Behaviour 36: 865-876.
Sandberg, R., Pettersson, J. & Alerstam, T. 1988b. Shifted magnetic fields lead to deflected and axial orientation of migrating Robins, Erithacus rubecula, at sunset. Animal Behaviour 36: 877-887.
Sandberg, R., Ottosson, U. & Pettersson, J. 1991. Magnetic orientation of migratory Wheatears (Oenanthe oenanthe) in Sweden and Greenland. Journal of Experimental Biology 155: 51-64.
Schmidt-Koenig, K. 1958. Experimentelle Einflußnahme auf die 24-Stunden-Periodik bei Brieftauben und deren Auswirkungen unter besonderer Berücksichtigung des Heimfindevermögens. Zeitschrift für Tierpsychologie 15: 301-331.
Schmidt-Koenig, K., Ganzhorn, J.U. & Ranvaud, R. 1991. The sun compass. In: Berthold, P. (ed.) Orientation in Birds. Basel; Birkhäuser Verlag: 1-15.
von Saint Paul, U. 1954. Nachweis der Sonnenorientierung bei nächtlich ziehenden Vögeln. Behaviour 6: 1-7.
Weindler, P. & Liepa, V. 1999. The influence of premigratory experience on the migratory orientation of birds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 979-987. Johannesburg: BirdLife South Africa.
Weindler, P., Beck, W., Liepa, V. & Wiltschko, W. 1995. Development of migratory orientation in Pied Flycatchers in different magnetic inclinations. Animal Behaviour 49: 227-234.
Weindler, P., Wiltschko, R. & Wiltschko, W. 1996. Magnetic information affects the stellar orientation of young bird migrants. Nature 383: 158-160.
Weindler, P., Böhme, F., Liepa, V. & Wiltschko, W. 1998. The role of daytime cues in the development of magnetic orientation in a night-migrating bird. Behavioral Ecology and Sociobiology 42: 289-294.
Wiltschko, R. 1980. Die Sonnenorientierung der Vögel. I. Die Rolle der Sonne im Orientierungssystem und die Funktionsweise des Sonnenkompaß. Journal für Ornithologie 121: 121-143.
Wiltschko, R. 1981. Die Sonnenorientierung der Vögel. II. Entwicklung des Sonnenkompaß und sein Stellenwert im Orientierungssystem. Journal für Ornithologie 122: 1-22.
Wiltschko, R. & Wiltschko, W. 1995. Magnetic orientation in animals. Berlin, Heidelberg, New York; Springer-Verlag: 297 pp.
Wiltschko, R., Kumpfmüller, R., Muth, R. & Wiltschko, W. 1994. Pigeon homing: the effect of clock-shift is often smaller than predicted. Behavioral Ecology and Sociobiology 35: 63-73.
Wiltschko, R., Munro, U., Ford, H. & Wiltschko, W. 1998. After-effects of exposure to conflicting celestial and magnetic cues at sunset in migratory Silvereyes, Zosterops l. lateralis. Journal of Avian Biology 30: in press.
Wiltschko, W. 1980. The relative importance and integration of different directional cues during ontogeny. In: Nöhring, R. (ed) Acta XVII Congressus Internationalis Ornithologici, Berlin 1978. Berlin; Deutsche Ornithologen-Gesellschaft: 501-565.
Wiltschko, W. & Wiltschko, R. 1975a. The interaction of stars and magnetic field in the orientation system of night migrating birds. I. Autumn experiments with European warblers (Gen. Sylvia). Zeitschrift für Tierpsychologie 37: 337-355.
Wiltschko, W. & Wiltschko, R. 1975b. The interaction of stars and magnetic field in the orientation system of night migrating birds. II. Spring experiments with European Robins ( Erithacus rubecula). Zeitschrift für Tierpsychologie 39: 265-282.
Wiltschko, W. & Wiltschko, R. 1991a. Magnetic orientation and celestial cues in migratory orientation. In: Berthold, P. (ed.) Orientation in Birds. Basel; Birkhäuser Verlag: 16-37.
Wiltschko, W. & Wiltschko, R. 1991b. Der Magnetkompaß als Komponente eines komplexen Richtungsorientierungssystems. Zoologisches Jahrbuch Physiologie 95: 437-446.
Wiltschko, W., Wiltschko, R. & Keeton, W.T. 1984. Effect of a "permanent" clock-shift on the orientation of experienced Homing Pigeons. Behavioral Ecology and Sociobiology 21: 333-342.
Wiltschko, W., Daum-Benz, P., Fergenbauer-Kimmel, A. & Wiltschko, W. 1987. The development of the star compass in Garden Warblers, Sylvia borin. Ethology 74: 285-292.
Wiltschko, W., Weindler, R. & Wiltschko, R. 1998a. Interaction of magnetic and celestial cues in the migratory orientation of passerines. Journal of Avian Biology 29: 606-617.
Wiltschko, W., Wiltschko, R., Munro, U. & Ford, H. 1998b. Magnetic versus celestial cues: cue-conflict experiments with migrating Silvereyes at dusk. Journal of Comparative Physiology 182: 521-529.
Table 1. Tests under the natural sky in deflected magnetic fields; individuals tested only once.
Table 2. Tests under the natural sky in deflected magnetic fields; individuals tested repeatedly.
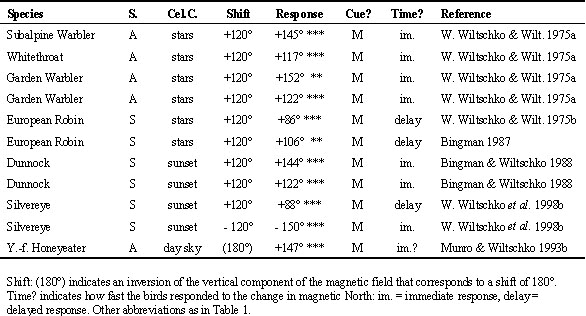
Table 3. Tests in the local geomagnetic field; manipulations of celestial cues.
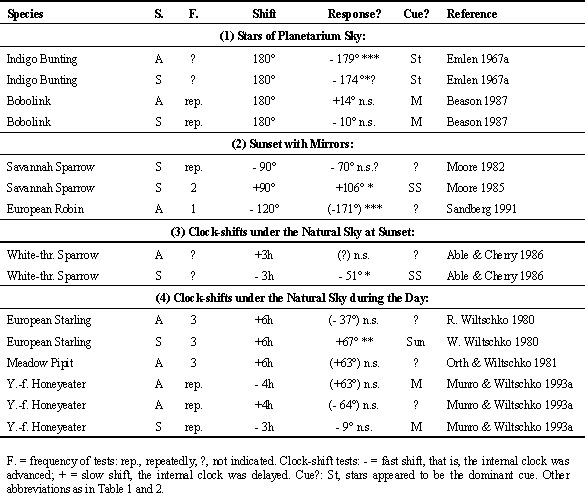
Table 4. After-effects of exposure to cue-conflict under the natural sky in deflected magnetic fields.
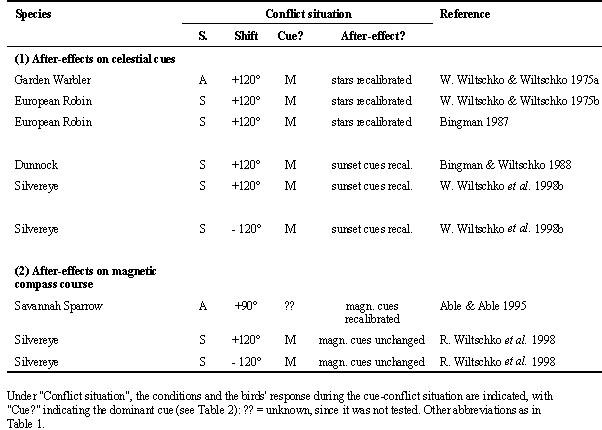
Fig. 1. Orientation of European Robins tested at night under the clear natural sky. Left: Birds tested in the local geomagnetic field; center and right: birds tested in a field deflected 120° clockwise to eastsoutheast; center: first two tests of each birds; right: third and later tests.- The symbols at the periphery of the circle give the nightly headings, the arrows represent the mean vectors with the lengths proportional to the radius of the circle = 1. The inner circles mark the 5% (dotted) and the 1% significance border of the Rayleigh test (data from Wiltschko & Wiltschko 1975b).
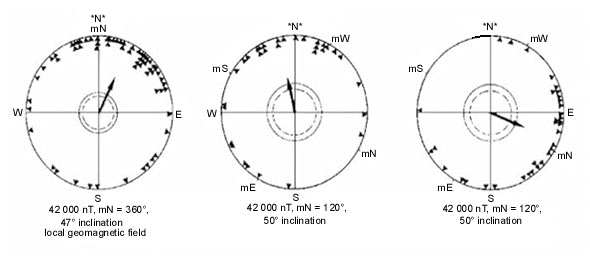
Fig. 2. Orientation of Australian Silvereyes in tests under conflicting celestial and magnetic cues and in subsequent tests with each set of cues alone. Left: Control birds; right: experimental birds. Upper diagrams: Tests under clear sky, control birds tested in the local geomagnetic field; experimental birds tested in a field deflected 120° counterclockwise to westsouthwest. Center diagrams: both groups subsequently tested under the clear sky without magnetic information. Lower diagrams: Both groups subsequently tested indoors with the magnetic field as only cue. - The symbols at the periphery of the circle give mean headings of individual birds; other symbols as in Fig. 1 (from R. Wiltschko et al. 1998).