S17.3: The influence of premigratory experience on the migratory orientation of birds
Peter Weindler 1 & Varis Liepa 2
1Zoologisches Institut, J. W. Goethe-Universität, Postfach 11 19 32, 60054 Frankfurt a. M., Germany, fax 49 69 79824820, e-mail weindler@zoology.uni-frankfurt.de; 2Biological Institute, Miera 3, 229021 Salaspils, Latvia
Weindler, P. & Liepa, V. 1999. The influence of premigratory experience on the migratory orientation of birds. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 979-987. Johannesburg: BirdLife South Africa.Pied Flycatchers, Ficedula hypoleuca, are known to use celestial and magnetic cues to orient their migratory flights. Studies investigating the ontogenetic relationship of these two cue systems revealed a complex interaction of celestial rotation and magnetic information on the stellar orientation of migrants. Celestial rotation provides information about a reference point only. This cue is modified by magnetic information into the population-specific migratory direction. These findings will be compared with those of similar experiments investigating conflicts of magnetic and celestial cues during the premigratory phase on the magnetic compass. For these experiments we used juvenile as well as adult Pied Flycatchers. Both groups of birds changed their magnetic compass course in response to celestial cues.
INTRODUCTION
Most night-migrating species migrate alone, and young birds and adults often migrate separately. This suggests that young birds possess sufficient inborn information that enables them to reach their wintering quarters. These observations were confirmed when hand-raised migrants were kept under controlled conditions in the laboratory. It was found that young birds possess genetically fixed information about the direction and distance of their migration route. This information is population-specific, i.e. when different populations of the same species follow different routes, like the western and eastern populations of Blackcaps, Sylvia atricapilla, in central Europe do, the genetically coded information on the migratory direction differs accordingly (Helbig & Wiltschko 1987). Genetically fixed directional information requires an external reference to be converted into an actual course of migration. Birds use two such reference systems, celestial rotation and the earth’s magnetic field.
Bird migrants can make use of the seemingly moving patterns of the night sky as well as during daytime to examine the axis of celestial rotation. In an elegant ontogeny experiment with Indigo Buntings, Passerina cyanea, Emlen (1970), with the help of a rotating planetarium sky, demonstrated the crucial role of rotation as a reference for the migratory direction. In autumn, Indigo Buntings headed away from the centre of rotation. Birds that had experienced a planetarium sky rotating around Betelgeuse in Orion instead of around the Polar Star readily accepted Betelgeuse as being in the north and oriented opposite to its azimuth. Information on celestial rotation is also provided by the changing pattern of polarised light during the day (Able & Able 1993; Weindler et al. 1998a) and at dawn and dusk (Able & Able 1995a). The use of the polarisation pattern during the day might be of particular importance for birds living at higher latitudes where the days are long during the premigratory period, with the stars visible only for a short period or not at all.
One effect of exposure to rotating stars during the premigratory period was that the birds later, during migration, were able to orient in their migratory direction under the same sky even when it was now stationary. This was true for a planetarium sky (Emlen 1970) as well as for an artificial sky with only 16 small lights as stars (W. Wiltschko et al. 1987; Weindler et al. 1996, 1997). This means that young birds use celestial rotation to determine their migratory course with respect to the stars, and, at the same time, establish a star compass that is, once functional, independent of celestial rotation. Thus, it appears that migrants, during migration, rely entirely on the star patterns they had learned during the premigratory period. The fact that the complex natural sky could be replaced by a simple artificial sky, provided the birds had experienced such a sky rotating with natural speed and direction, clearly shows that the composition of the sky itself is irrelevant. Birds do not seem to have innate knowledge of the appearance of the sky, as was once suggested (Sauer 1957), but can learn to use any stars. The crucial cue for establishing a star compass is the rotation of the stars around a celestial pole.
While celestial rotation is a highly important cue, it does not seem to be sufficient for the complete transformation of the genetically coded information on the migration course. Garden Warblers, Sylvia borin, of the central European population start migration by heading about 225° southwest (Zink 1985), i.e. their migratory direction deviates markedly from south. During the premigratory period, Garden Warblers were exposed to a rotating artificial sky in the absence of magnetic information. When the birds were later tested with the same, now stationary stars as the only available cue, they headed away from the centre of rotation, a direction that corresponds to stellar south instead of the population specific southwesterly course (Weindler et al. 1996). Similar tendencies towards due south were also observed in Garden Warblers that had been exposed to an artificial sky rotating in the reversed direction (Weindler et al. 1997). These findings indicate that celestial rotation provides only a reference direction away from the centre of rotation. This direction corresponds to geographic south and would lead the birds towards the equator. In this situation the direction of rotation, either clockwise or counterclockwise, would appear to be irrelevant. The population-specific southwesterly orientation, however, was observed only in Garden Warblers that had experienced celestial rotation together with the geomagnetic field.
Thus, the magnetic field of the earth must provide the second reference for the innate information on the migratory course. Young birds of several species that had been hand-raised without access to celestial cues (for Pied Flycatchers see Beck & Wiltschko 1982) were able to orient in the appropriate direction during autumn migration with the geomagnetic field as the only available cue (for summary see R. Wiltschko & Wiltschko 1995). Birds with migratory directions deviating markedly from due south mostly showed their specific southwesterly or southeasterly courses. The resulting non-straight migration routes can be closely adapted to the specific situation of each population and allow birds to largely avoid ecological barriers. Central European Garden Warblers and Pied Flycatchers thus bypass the Alps and the Mediterranean Sea and cross the Sahara at its western edge where less harsh conditions prevail.
The finding that both celestial rotation and the magnetic cues serve as a reverence for the migratory course raised the question about a hierarchy of cues and possible interactions between the two systems. This question was approached by exposing hand-raised young birds during the premigratory period to the natural or an artificial rotating sky in deflected magnetic fields. The angular deviation between rotational and magnetic north was experimentally turned between 90 and 140 degrees. Later during autumn migration, these birds were tested either with the magnetic field as the only cue in order to look for a possible effect of celestial information on the magnetic compass course, or with celestial cues in the absence of magnetic information to look for an effect of magnetic information on the stellar course.
The results of these experiments demonstrate that exposure to a cue-conflict situation of celestial and magnetic cues resulted in an altered magnetic compass course. The birds did no longer prefer their normal migratory direction with respect to the magnetic field, but oriented along that magnetic direction that was indicated by celestial rotation. This shows that in case of cue-conflict between rotational (geographic) north and magnetic north, the birds chose celestial rotation over the magnetic field and adjusted their magnetic compass course accordingly. While tests with the magnetic field as the only cue revealed a strong influence of celestial rotation on the magnetic compass course, the opposite type of tests with stars as the only cue showed that during the premigratory period, the magnetic field had little effect on the compass course with respect to the stars. Although the difference between rotational north and magnetic north was more than 100°, the behaviour of Pied Flycatchers under the natural night sky and of Garden Warblers under an artificial sky did not differ from that of control birds that had been exposed to celestial rotation and magnetic north in their normal relationship. In these cases, the birds headed in the direction away from the centre of rotation (Bingman 1984; W. Wiltschko et al. 1987).
Available experimental evidence clearly shows that during the premigratory period, cues provided by celestial rotation dominate cues provided by the magnetic field. The question remains open whether this is true not only for juvenile migrants on their first autumn migration but also for adult birds that have travelled at least one full migration cycle. Therefore the aim of this study is to compare the orientation behaviour of juvenile and adult Pied Flycatchers during autumn migration with the magnetic field as the only cue.
METHODS
The tests took place at the Ornithological Research Station of the Latvian Institute of Biology near Tukums (56°55'N, 23°10'E), about 60 km west of Riga, Latvia. The local geomagnetic field has an intensity of 50 000 nT, with 73° inclination.
Test Birds
The test birds were Pied Flycatchers, long-distance migrants, that migrate mostly at night. In autumn, they leave their breeding areas in central and northern Europe heading southwest with a course of about 225°, as indicated by ringing recoveries. Later in October, after passing the Iberian peninsula, they shift to a more southerly course towards their wintering areas in western Africa south of the Sahara (Zink 1985). In previous studies, Pied Flycatchers in Latvia had been shown to need access to celestial rotation for unimodal orientation in their migratory direction (Weindler et al. 1995, 1998a, b).
Juvenile test birds were taken from their nest-boxes near the Ornithological Research Station at the age of about 10 days. They were transferred to the laboratory of the station and hand-raised until they became self-sufficient. Rearing conditions were similar for all birds (for details, see Weindler et al. 1995). Adult birds were captured from the same nestboxes as their young.
Exposure during the Premigratory Period
All birds lived in aviaries in a windowless room under an artificial light regime that corresponded to the local photoperiod in the natural local geomagnetic field. They were subdivided into four groups that differed with respect to age and to exposure to celestial and magnetic cues during the premigratory period.
Group AC is the control group of adult individuals. They were exposed under the natural outdoor conditions, where they experienced natural rotation of the sky and magnetic north coinciding.
Group AE is the experimental group of adult individuals. They were exposed to the natural outside sky with magnetic north turned by 90° to geographic west with the use of a Helmholtz coil.
Group JC is the control group of juvenile individuals. They were exposed under a planetarium sky simulating the natural conditions. This group corresponds to the LST birds published in Weindler et al. (1995).
Group JE is the experimental group of juvenile individuals. They were exposed under a planetarium sky with rotational north shifted by 140° to magnetic SE.
The exposure lasted for about 4 weeks; after that period, none of the birds was allowed access to visual cues.
Orientation Tests
Orientation tests were performed during August and September, i.e. during the first part of autumn migration when Pied Flycatchers prefer southwesterly headings. Tests took place inside wooden huts where the birds experienced the local geomagnetic field but had no access to celestial cues. The test rooms were lit by small light bulbs the light of which passed through diffusing Plexiglass.
The birds' activity was recorded using funnel cages (Emlen & Emlen 1966) lined with typewriter correction paper (Tipp-Ex, Germany). By their movements, the birds left scratches on the coating of the inclined walls which could later be counted. For details of the test arrangement, see Weindler et al. (1995).
Data Analysis and Statistics
The Tipp-Ex paper recordings were divided into 24 sectors of 15°, and the scratches were counted on a light table. From their distribution, the heading of the respective test was calculated by vector addition. Tests with fewer than 40 scratches were not considered because of too little activity. From the headings of each individual bird, the mean vector was calculated. The Rayleigh-test (Batschelet 1981) was used to test for directional preference. The Mardia-Watson-Wheeler-test and the Watson-Williams-test were used to test for statistical differences between samples.
RESULTS
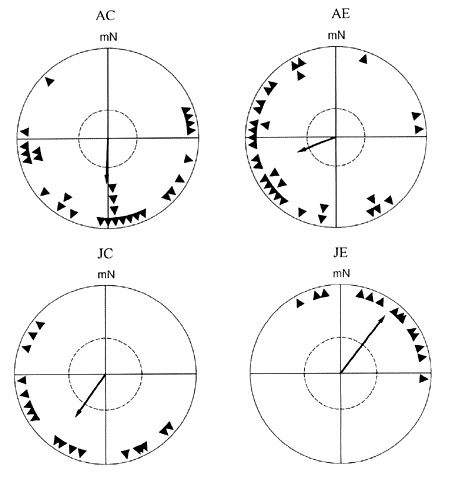
The orientation behaviour during autumn migration of juvenile as well as of adult Pied Flycatchers is given in Fig. 1. There is a significant difference in the orientation behaviour of juvenile birds between the control group and the experimental group (Mardia-Watson-Wheeler-test, P < 0.001). The control birds that were exposed to a planetarium sky, simulating the natural conditions, oriented during autumn migration in a southerly direction (n = 18, a = 215°, r = 0.59, P < 0.01), whereas birds raised under a cue-conflict situation with magnetic and celestial north differing by about 140° oriented into a north-easterly direction (n = 14, a = 37°, r = 0.82, P < 0.001).
Adult Pied Flycatchers that had been exposed under natural conditions during the premigratory period, showed a southeasterly or southwesterly mean direction which do not differ significantly (1993: n = 8, a = 144°, r = 0.50, P = 0.06; 1995: n = 21, a = 197°, r = 52, P < 0.01). Adult birds that had experienced the natural sky in a shifted magnetic field where magnetic north coincided with geographic west oriented into a westerly direction (1993: n = 8, a = 230°, r = 0.76, P < 0.01; 1995: n = 23, a = 262°, r = 0.39, P < 0.05). Thus in both years the experimental group oriented into a more westerly direction that differs from that of the control group about +86° in 1993 and about +65° in 1995 respectively. The differences between the control group and the experimental group are not significant within each year. However, if both years are taken together the experimental mean direction (n = 31, a = 249°, r = 0.46, P < 0.01) differs significantly (Mardia-Watson-Wheeler-test, P < 0.05; Watson-Williams-test, P < 0.01) from the direction of the control group (n = 29, a = 182°, r = 0.49, P < 0.001).
DISCUSSION
The results of our experiment clearly demonstrate that adult as well as juvenile Pied Flycatchers adjust their magnetic compass course in response to celestial cues. When exposed during the premigratory period to a cue-conflict situation between celestial and magnetic cues, the birds changed their migratory direction relative to the control groups that had experienced celestial and magnetic cues under natural conditions with magnetic and geographic north nearly coinciding (-1.5° declination). Our results confirm findings by Able & Able (1995b), demonstrating that adult and juvenile Savannah Sparrows, Passerculus sandwichensis, reacted in a similar way when exposed to a cue-conflict situation during autumn migration.
At the first glance the adjustment of the magnetic compass course might be important only in areas where birds are confronted with rather large deviations between the magnetic and the geographic pole as it is the case, for example, in the northern regions of America. However, the degree of the magnetic deviation is not only flexible in space but also in time. At London, the declination changed over the last three centuries by about 30°. Thus, in evolutionary time migrating species were confronted with the problem of changing declinations. It seems that migrants therefore developed a mechanism to adjust their magnetic compass course in relation to geographic north.
The interaction between celestial cues and the geomagnetic field is more complex than was first thought. The adjustment of the magnetic compass course does not work by simply overwriting the magnetic cues by celestial rotation. Studies with Garden Warblers indicate that celestial rotation per se provides information about an axis only, like away from the centre of rotation, corresponding to geographic south (Weindler et al. 1996), whereas the population-specific southwesterly course seems to be encoded with respect to the magnetic field (R. Wiltschko & Wiltschko 1995). Birds could orient in their seasonally appropriate southwesterly direction by stationary artificial stars alone, as long as they had observed these stars rotating while being exposed to the geomagnetic field (W. Wiltschko et al. 1987; Weindler et al. 1996, 1997). Apparently, information from celestial cues has to be combined with information from the magnetic field to establish the population-specific migratory direction with respect to the stars.
Even when the magnetic compass course was altered by exposing birds to celestial rotation in deflected magnetic fields, species that normally head magnetically southwest preferred the magnetic direction that had corresponded to geographic southwest during exposure and not the one that had corresponded to south as directly away from the centre of rotation (Weindler & Wiltschko 1991; Prinz & Wiltschko 1992). It means that even when the magnetic compass course is overruled by celestial rotation, information encoded with respect to the magnetic field is integrated in establishing the migratory course. This seeming paradox can be best explained by the assumption that the crucial information on the population-specific course is not encoded as a certain magnetic course, but as a deviation from a reference direction. In the case of the Garden Warbler, that would mean that the innate information does not specify ‘225° SW’ with respect to the magnetic field, but something equivalent to ‘45° clockwise’ of a reference.
This reference can be provided by both, celestial rotation and the geomagnetic field, corresponding to geographic south and magnetic south (equatorwards), respectively. The dominance of celestial rotation over the magnetic field suggests that birds normally use celestial rotation. Yet, when there is no access to celestial information, magnetic south can be used instead; that is clearly demonstrated by the oriented behaviour of birds that never saw the sky (for summary see R. Wiltschko & Wiltschko 1995).
For any interaction between celestial and magnetic cues, the direction of celestial rotation proved crucial. Garden Warblers that had experienced a rotating artificial sky together with the magnetic field preferred their normal southwesterly course when the sky had rotated in the direction appropriate for the northern hemisphere: counterclockwise around the polar star. Exposed under a sky moving in the reversed direction, they later headed due south (Weindler 1997; Weindler et al. 1997). Their behaviour indicates that with reversed rotation, the information on the deviation from the reference direction coded with respect to the magnetic field could not be combined with the reference given by celestial rotation. As a result, the birds had to rely on the information provided by celestial rotation alone, namely away from the centre of rotation.
For the adjustment of the magnetic compass course the direction of rotation also proved crucial. When hand-raised Pied Flycatchers were exposed to the natural daylight or to a planetarium sky where the stars moved in the natural direction, the birds were able to combine celestial and magnetic cues (Weindler et al. 1995, 1998a), whereas birds exposed to a planetarium sky where the stars were rotating in the reversed direction could not (Weindler et al. 1998b). Together, these findings point out a crucial role of the direction of celestial rotation when magnetic and celestial information are combined.
Thus in nature, celestial and magnetic cues interact during the premigratory period to convert genetically fixed information into the starting course of migration, celestial rotation normally providing the reference direction, and the magnetic field providing the population-specific deviation from that reference. Once that course is established, it can be located with the help of the star compass, the magnetic compass and also with the sunset-related cues such as the setting sun and the typical pattern of polarised light at sunset.
During migration, however, the dominance of celestial rotation seems to disappear whereas magnetic cues gain dominance. The analysis of experiments conducted with mistnetted birds during actual migration documents a completely different picture than that observed before the onset of migration (for summary see Akesson 1994; R. Wiltschko et al. 1997). The reason why the relative importance of celestial and magnetic cues change during the season is unclear, yet.
Possible reasons may lie in the nature of the cues and their geographic variation. As birds migrate, they usually move from higher to lower latitudes. Features of the magnetic field that might restrict its usefulness as a reference for innate directional information, such as large and fast-changing magnetic declinations, quickly disappear as the birds reach temperate latitudes. The magnetic field becomes an increasingly regular, reliable factor that is available irrespective of weather conditions. Celestial rotation, which has been the crucial aspect of celestial cues during the premigratory period, ceases to play a role, as the tests under stationary stars indicate (Emlen 1970; W. Wiltschko et al. 1987). Another possibility is that birds can no longer extract information from rotation when they are on the move.
The ability to perceive information from celestial cues via celestial rotation seems to be restricted to juvenile birds within a rather short time period between fledging and the onset of autumn migration only. Emlen (1970) revealed that young birds have to locate the region of the stellar sky where the angular velocities of star rotation are minimal. Thus the centre of celestial rotation represents a key stimulus which activates a genetically pre-programmed response. How adult birds adjust their magnetic compass course with the help of celestial cues remains unclear.
Existing experimental evidence from birds tested during migration show that celestial cues are calibrated by the magnetic field. This seems to be true for the stars (W. Wiltschko & Wiltschko 1975a, b, 1991) and for the patterns of polarised sunlight (Able & Able 1990) as well as for the sun (R. Wiltschko 1983; W. Wiltschko & Wiltschko 1988). The most plausible explanation seems to be that for juvenile as well as for adult birds there is a sensitive period before each migratory season starts. The open question is whether adult birds are able to perceive information from the rotation of celestial cues or not (Liepa 1994). If not, the adjustment of the magnetic compass course, as we could observe it in our experiment, might work in a different way. Adult birds then have to remember the stellar configurations they learned during their juvenile premigratory period. Stars around the former centre of rotation then could be used to compare with the actual magnetic direction and to set the new magnetic compass course, respectively. However, the direct use of rotational information has never been studied. Searching for experimental methods in another biological context to test the role of this information in the orientation system of birds should be the subject of future investigations.
ACKNOWLEDGEMENTS
Our research was supported by the Deutsche Forschungsgemeinschaft, the Deutsche Ornithologen-Gesellschaft and the Freunde und Förderer der J.W.Goethe-Universität Frankfurt a.M. as well as by stipends of the Deutscher Akademischer Austauschdienst and of the Graduiertenförderung des Landes Hessen to P.W.. We thank G. Strauts for his help in setting up the Latvian test arrangement and F. Böhme for his help with conducting the experiments.
REFERENCES
Able, K.P. & Able, M.A. 1990. Calibration of the magnetic compass of a migratory bird by celestial rotation. Nature 347: 378-389.
Able, K.P. & Able, M.A. 1993. Daytime calibration of magnetic orientation in a migratory bird requires a view of skylight polarization. Nature 364: 523-525.
Able, K.P. & Able, M.A. 1995a. Manipulations of the polarized skylight calibrate magnetic orientation in a migratory bird. Journal of Comparative Physiology 177: 351-356.
Able, K.P. & Able, M.A. 1995b. Interaction in the flexible orientation system of a migratory bird. Nature 375: 230-232.
Akesson, S. 1994. Comparative orientation experiments with different species of passerine long-distance migrants: effect of magnetic field manipulations. Animal Behaviour 48: 1379-1393.
Batschelet, E. 1981. Circular Statistics in Biology. New York; Akademic Press.
Beck, W. & Wiltschko, W. 1982. The magnetic field as reference system for the genetically encoded migratory direction in Pied Flycatchers. Zeitschrift für Tierpsychologie 60: 41-46.
Bingman, V.P. 1984. Night sky orientation of migratory Pied Flycatchers raised in different magnetic fields. Behavioural Ecology and Sociobiology 15: 77-80.
Emlen, S.T. 1970. Celestial rotation: its importance in the development of migratory orientation. Science 170: 1198-1201.
Emlen, S.T. & Emlen, J.T. 1966. A technique for recording migratory orientation of captive birds. The Auk 83: 361-367.
Helbig, A. & Wiltschko, W. 1987. Untersuchung populationsspezifischer Zugrichtungen der Mönchsgrasmücke Sylvia atricapilla mittels der Emlen-Methode. Journal für Ornithologie 128: 311-316.
Liepa, V. 1994. Do birds respond to celestial rotation indeed only before the onset of first migration? Ring 16: 16-21.
Prinz, K. & Wiltschko, W. 1992. Migratory orientation of Pied Flycatchers: interaction of stellar and magnetic information during ontogeny. Animal Behaviour 44: 539-545.
Sauer, F. 1957. Die Sternorientierung nächtlich ziehender Grasmücken, Sylvia atricapilla, borin und curruca. Zeitschrift für Tierpsychologie 14: 20-70.
Weindler, P. 1997. Celestial rotation - information about an axis only? In: Orientation and Navigation - Birds, Humans and Other Animals. Oxford; Royal Institute of Navigation: 7-1: 7-9.
Weindler, P. & Wiltschko, W. 1991. Angeborene und erlernte Verhaltensmechanismen junger Gartengrasmücken, Sylvia borin auf ihrem ersten Zug. Verhandlungen der Deutschen Zoologischen Gesellschaft 84: 369.
Weindler, P., Beck, W., Liepa, V. & Wiltschko, W. 1995. Development of migratory orientation in Pied Flycatchers in different magnetic inclinations. Animal Behaviour 49: 227-234.
Weindler, P., Wiltschko, R. & Wiltschko, W. 1996. Magnetic information affects the stellar orientation of young bird migrants. Nature 383: 158-160.
Weindler, P., Baumetz, M. & Wiltschko, W. 1997. The direction of celestial rotation influences the development of stellar orientation in young Garden Warblers. Journal of Experimental Biology 200: 2107-2113.
Weindler, P., Böhme, F., Liepa, V. & Wiltschko, W. 1998a. The role of daytime cues in the development of magnetic orientation in a night-migrating bird. Behavioral Ecology and Sociobiology 42: 289-294.
Weindler, P., Liepa, V. & Wiltschko, W. 1998b. The direction of celestial rotation affects the development of migratory orientation in Pied Flycatchers. Ethology: in press.
Wiltschko, R. 1983. The ontogeny of orientation in young pigeons. Comparative Biochemistry and Physiology 76A: 701-708.
Wiltschko, R. & Wiltschko, W. 1995. Magnetic orientation in animals. Berlin Heidelberg; Springer-Verlag: 297pp.
Wiltschko, R., Wiltschko, W. & Munro, U. 1997. Migratory orientation in birds: the effects and after-effects of exposure to conflicting celestial and magnetic cues. In: Orientation and Navigation - Birds, Humans and Other Animals. Oxford; Royal Institute of Navigation: 6-1 - 6-14.
Wiltschko, W. & Wiltschko, R. 1975a. The interaction of stars and magnetic field in the orientation system of night migrating birds. I. Autumn experiments with European Warblers (Gen. Sylvia). Zeitschrift für Tierpsychology 37: 337-355.
Wiltschko, W. & Wiltschko, R. 1975b. The interaction of stars and magnetic field in the orientation system of night migrating birds. II. Spring experiments with European Robins, Erithacus rubecula. Zeitschrift für Tierpsychology 39: 265-282.
Wiltschko, W. & Wiltschko, R. 1988. Magnetic orientation in birds. In: Johnston R.F. (ed) Current Ornithology. Vol 5; New York; Plenum Press: 67-121.
Wiltschko, W. & Wiltschko, R. 1991. Magnetic orientation and celestial cues in migratory orientation. In: Berthold, P. (ed), Orientation in birds. Basel; Birkhäuser: 16-37.
Wiltschko, W., Daum-Benz, P., Fergenbauer-Kimmel, A. & Wiltschko, R. 1987. The development of the star compass in Garden Warblers. Ethology 74: 285-292.
Zink, G. 1985. Der Zug europäischer Singvögel. Ein Atlas der Wiederfunde beringter Vögel. 4. Lieferung; Möggingen; Vogelzug-Verlag.
Fig. 1. Orientation of Pied Flycatchers in the local geomagnetic field during autumn migration. The headings of the birds are represented by solid dots at the periphery of a unit circle = 1. Mean vectors are represented by arrows. The inner circle (broken line) indicates the 5% significance level of the Rayleigh test. mN = magnetic north. AC and AE are adult birds. AC = control group, exposed during the premigratory period to natural conditions. AE = experimental group, exposed outside, where magnetic north was shifted -90° to geographic west. JC and JE are juvenile birds. During the premigratory period they were exposed under a planetarium sky. For group JC the natural situation was simulated, whereas for group JE the centre of rotation was shifted by 140° to geographic SE.