S17.1: How do young night migrating passerines use their compasses for orientation during migration?
Henrik Mouritsen
Centre for Sound Communication, Institute of Biology, Odense University, Campusvej 55, DK-5230 Odense M, Denmark, fax 1 613 533 2499, e-mail
mou@ psyc.queensu.ca *Present address: Department of Psychology,Queen's University, Kingston, Ontario K7L3N6, Canada. Mouritsen, H. 1999. How do young night migrating passerines use their compasses for orientation during migration? In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 934-955. Johannesburg: BirdLife South Africa.Are night migrating passerines during their first migration simple clock-and-compass machines just flying in a set species-specific compass direction, or are they actively navigating? Recently, I developed a mathematical model, which showed that the directional distributions as a function of distance found in ringing recoveries of Scandinavian Pied Flycatchers, Ficedula hypoleuca, correlated strongly with those predicted from the model of the simple clock-and-compass strategy. As an empirical test, a displacement experiment was performed. Pied Flycatchers caught in Scandinavia during their first autumn migration were tested before and after being displaced about the maximal biologically realistic distance due south and due west. The birds showed no signs of compensatory orientation. Also, I recently reviewed the displacement literature and found that the evidence supporting the clock-and-compass hypothesis strongly outweighed the evidence against it. All this evidence suggests that young night migrating passerines are equipped with compasses only, but no map.
INTRODUCTION
Each autumn, millions of night-migrating passerine birds leave their breeding grounds in northern Europe and migrate several thousand kilometres to their winter quarters in tropical Africa (for a review see Moreau 1972). More than 50% of these birds are in their first year and must find the unfamiliar winter quarter on their own, without guidance from experienced adults. Therefore, they must rely exclusively on inherited orientation strategies.
At present, it is well established that night migrating passerines have an inherent time schedule (for a review see Berthold 1991; Gwinner 1996), and that they possess at least two compasses, a magnetic compass and a celestial compass (e.g. Moore 1987; Alerstam 1990; W. Wiltschko & Wiltschko 1991, 1996; Mouritsen 1998), and that an at least initial migratory direction is inherited (for a review see Helbig 1996). To find their species (population) specific winter quarters, night migrating passerines need to integrate the time and spatial information from these prerequisites into a spatiotemporal orientation programme.
However, the question on how these prerequisites are used has been a subject of some controversy (see e.g. Gwinner & Wiltschko 1978; R. Wiltschko & Wiltschko 1978; Rabøl 1978, 1981, 1994; Berthold 1991; Alerstam 1996). Two main views exist: Young night migrating passerines use a clock-and-compass strategy or they use true navigation. In other words, do night migrating passerines have both compasses and a map or compasses only?
The clock-and-compass model (a vector navigation hypothesis, see e.g. Berthold 1991, pp. 97-99; see also Beck 1984) describes the migratory programme as a series of unrelated migratory steps oriented in an almost constant direction (the compass) under a predetermined time schedule (the clock). The clock component controls when to start and stop migration and may trigger changes in the preferred compass direction once or twice during one migratory season. According to the clock-and-compass model, the birds do not need to possess any map-sense, and they are not able to navigate and thereby correct for directional mistakes (except perhaps via emergency plans, see later). Given the compass abilities of birds, the compass component of the clock-and-compass mechanism is the most simple spatiotemporal orientation mechanism one can think of. But importantly, night migrating passerines are known to have the time schedule and the compasses needed for such a mechanism to work.
The ‘Co-ordinate’ or Goal Area Navigation hypothesis (see e.g. Rabøl 1969, 1978, 1994) suggests that the orientation programme of night migrating passerines is a true navigational process in which the birds continuously orient towards a moving goal area or the wintering area. A moving goal area is the optimal geographical place to be at any particular time during migration (Rabøl 1978). Such a goal area will move along the normal migratory route as time proceeds. A true navigational system would enable birds to correct for directional mistakes. In order to perform true navigation, birds need both an inherited compass and a map. Here, a `map' should be understood in its broader sense. The `map' may be very simple indeed, perhaps consisting only of inherited information about the co-ordinate set (functioning like longitudes and latitudes) of the wintering area and/or of a few fix points along the migration route. Such a strategy would be fundamentally different from a vector navigation strategy since the compass would not just be used for setting one constant innate direction, but for setting different directions depending on position relative to e.g. wintering area co-ordinates. The fact that animals can inherit a few co-ordinate sets, which forms a simple `map', is known from hatchling sea turtles (see e.g. Lohmann & Lohmann 1996). They orient in different directions when presented with different combinations of magnetic strength and inclination. This behaviour ensures that the young sea turtles stay within the North Atlantic Gyre.
In short, most students of bird migration tend to support the clock-and-compass hypothesis. However, in relation to night migrating passerines this support has previously been almost entirely based on evidence concerning the clock-component (for reviews see Berthold 1991; Gwinner 1996) but has largely ignored that (numerous) studies on the compass-component (all conducted by Dr Jørgen Rabøl) provided conflicting evidence that mainly was interpreted as supporting the navigation hypothesis. By the compass component, I do not mean the compasses themselves (they are well-described), but how they are used over time during migration.
THE CLOCK/TIME-COMPONENT
The following points of special relevance to our understanding of the role of circannual clocks in the endogenous control of migration can be extracted from the reviews of Berthold (1991) and Gwinner (1996).
First, there is fairly good accordance between onset, duration and termination of migratory restlessness in caged migrants and the course of actual migration in wild conspecifics (e.g. Berthold 1975).
Second, comparative studies in groups of warblers of the genera Phylloscopus and Sylvia (using species and populations with different migratory performances) have demonstrated that long-distance migrants show a high degree of migratory restlessness, middle-distance migrants lower and short distance migrants the least (e.g. Berthold 1973; Gwinner 1986).
Third, Berthold & Querner (1988) calculated that the duration of migratory restlessness expressed through wing whirring in caged Garden Warblers, Sylvia borin, multiplied by the known species specific flight speed during migration would have brought the experimental group into the centre of the species specific winter quarters. Thus, migratory restlessness may be interpreted (although with a number of reservations) as migration by wing whirring in a sitting (or jumping) position.
Fourth, Sauer (1957) tested the directional preferences of a Lesser White-throat, Sylvia curruca, (‘Müllerchen’) under several star constellations mimicking different latitudes along its migratory route. The Lesser White-throat showed south-easterly and southerly headings, respectively, when presented with latitudes higher and lower than 30° N. This led Sauer to claim that Lesser White-throats use the star constellations for navigation and that they trigger the directional shift from south-east to south observed in freely migrating Lesser White-throats at latitudes of approximately 30° N. It is important to note, however, that these general conclusions were based mainly on several experiments on one individual only. Also, Emlen attempted to repeat Sauer's experiments but failed to confirm his results (A. Helbig, personal communication).
Fifth, in 1978, Gwinner & Wiltschko found that Garden Warblers tested continuously in southern Germany without access to celestial cues during August to December spontaneously showed the species specific change in orientation from south-west to south-south-east at the time when their free-living conspecifics do so. Similar results have also been found for Yellow-faced Honeyeaters, Lichenostomus chrysops, (Munro et al. 1993) and Blackcaps, Sylvia atricapilla, (Helbig et al. 1989). Consequently, they suggested that the directional shift observed in Garden Warblers and many other bird species may be controlled by the endogenous time schedule alone. This suggestion is supported by an experiment by Gwinner (1968), in which he transported Willow Warblers, Phylloscopus trochilus, to their wintering areas and allowed them to see the local sky conditions. This did not stop their migratory activity, since they continued to show migratory restlessness for as long as conspecific controls, which were tested in the breeding area. Unfortunately, Gwinner (1968) did not test the directional preferences of the displaced or control birds. Gwinner & Wiltschko (1978) also re-analysed the data of Sauer (1957), and found that the reported directional shift was equally well correlated with the time of year as with the star constellations presented to the birds. Thus, Sauer’s data could be the result of either an endogenous time control or from the changed star constellations. However, as pointed out by Gwinner & Wiltschko (1978), none of the existing data on the time component can be used to distinguish between the clock-and-compass hypothesis and the goal area navigation hypothesis.
In conclusion, the clock/time component of the spatiotemporal orientation programme of night migrating passerines may provide the distance component required for a simple clock-and-compass strategy or set the time frame for navigation.
THE COMPASS COMPONENT
Perdeck’s (1958) impressive displacement experiment with European Starlings, Sturnus vulgaris, is the classical textbook experiment, which strongly suggests that juvenile birds use a clock-and-compass mechanism for orientation. Thousands of birds were ringed in the Netherlands in autumn. About half of these were displaced to Switzerland and released. The rest were released at the site of capture. An analysis of the recoveries of birds from the two groups from the same autumn and winter revealed a striking and highly significant difference between adult and juvenile birds. The controls ringed and released in the Netherlands all oriented in the normal south-westerly migratory direction and both the adults and the juveniles ended up in northern France. The displaced juvenile starlings continued the same distance as the control birds on their usual south-westerly course and ended up in southern France and Spain, whereas the displaced adult starlings corrected for the displacement and ended up close to or in the normal wintering quarter in northern France. Perdeck’s experiment provides convincing evidence that young European Starlings use a simple clock-and-compass strategy if left on their own, whereas adult European Starlings seem to have acquired navigational abilities through associative learning and experience.
Can Perdeck’s experiment be generalised to all birds, including night migrating passerines? European Starlings migrate relatively short distances mainly during the day and most importantly they migrate in large flocks. This means that all a young European Starling theoretically needs to do on its first migration is to stick close to its parents or other adults and learn where to go en route the first time. Therefore, the clock-and-compass mechanism demonstrated by Perdeck might function as a crude emergency plan activated only if a young bird finds itself deserted by adults. In comparison, young night migrating passerines have to migrate much longer distances at night and most importantly they do so alone without guidance from adult individuals. Therefore, selection may have forced them to use a more sophisticated orientation mechanism. Consequently, Perdeck’s demonstration that young European Starlings use a simple clock-and-compass mechanism cannot be generalised to night migrating passerines (or any other type of birds) until experiments on such birds have rendered its use probable.
Experiments in Emlen funnels (see e.g. Emlen & Emlen 1966; Mouritsen & Larsen 1998) before and after geographical displacement of night migrating passerines caught during their first migration has been used to study the nature of the directional component of their inherited orientation programmes (see e.g. Rabøl 1969, 1978, 1994). If birds do not compensate for geographical displacements, a ‘clock-and-compass’ programme would be suggested. If they compensate (i.e. turn towards their normal migratory route) several explanations should be considered. These include goal-area navigation, ‘co-ordinate’ navigation, path-integration (i.e. the birds detect the direction of displacement during transport and establish a reverse vector component by means of a compass) and cross-axis navigation (a simple sort of sign navigation). An excellent description of these is given by Rabøl (1994). In addition, it could theoretically be important that the birds undergo a fast shift of ½-1 hour when displaced east-west (W. Wiltschko, personal communication). The birds may react with a tendency to go to the left by a few degrees. There is indication in the literature that migrants react to such clock-shift around sunset, even when the sun is not directly visible (Able & Cherry 1986; Helbig 1991). Despite numerous experiments, the results remain unclear.
To dig deeper into the problem of how night migrating passerines make use of their compasses during migration, I studied the use of their compass component in their spatiotemporal orientation programmes from three different angles making up the classic tripod consisting of theory, lab experiments and field data (the real world). First, a mathematical model of clock-and-compass migration was developed (Mouritsen 1998b). Second, the model predictions were compared with data from the real world (Mouritsen 1998b). Third, the model predictions were tested in field laboratory experiments (Mouritsen & Larsen 1998). Fourth, the literature was reviewed and re-evaluated.
The mathematical modelIf a night migrating passerine follows a simple clock-and-compass strategy, its migration can be described as a series of unrelated random choices from a circular normal-like distribution. In other words, if a bird for example makes a large mistake to the right of the average migratory direction during any migratory step, the probability of going left of the average direction during the following migratory steps is not increased. Thus, birds using a clock-and-compass strategy will accumulate errors whereas birds with navigational abilities will be able to correct for errors. This means that the ringing recoveries would be expected to show a higher directional concentration if birds possess navigational abilities than if they use a simple clock-and-compass strategy only. Therefore, a mathematical model was made (Mouritsen 1998b), which predicted the expected directional distribution as a function of distance covered and of the birds’ ability to fly in the migratory direction under the assumption that they are using a simple clock-and-compass strategy (see Fig. 1).
Comparing model predictions with data from natural migrationThe model becomes relevant to our understanding of natural migration only if its predictions can be compared to data from naturally migrating birds. Pied Flycatchers, Ficedula hypoleuca, from the Scandinavian population were chosen as the ideal species for such comparisons. First, young Pied Flycatchers, as night migrating passerines in general, migrate alone without guidance from their parents. Second, all Pied Flycatchers migrate towards western Africa and no migratory divides exist (Zink 1985). This eliminates the risk of including birds from another population using a different migration route, which could contaminate the results. Third, huge numbers of Pied Flycatchers have been ringed in Scandinavia. This means that a large number of ringing recoveries is available. Fourth, most Pied Flycatchers ringed in Scandinavia were banded in nestboxes making the ringing sites widely dispersed geographically. This reduces the sources of error when analysing the recovery data.
To compare predictions from the model with ringing recovery data, Pied Flycatchers’ ability/precision to fly in the migratory direction had to be estimated. This ability can be estimated by studying ringing recoveries of birds found at a distance of approximately one migratory step from the banding site within the same autumn (for definition of a same autumn recovery, see Mouritsen 1998b).
One migratory step is likely to be in the order of 125 km in northern Europe (see Mouritsen 1998b). Therefore, I calculated the directional concentration (expressed by the length of the sample mean vector (the higher the r-value the larger the concentration), see Batschelet 1981) of birds recovered at distances of 100 to 149 km from the banding sites (see Fig. 2). It is important to note that the ringing recovery data came from birds being banded at geographical locations widely scattered across Scandinavia over a time span of more than 50 years. By basing the estimate of birds’ ability to fly in the migratory direction on such data, naturally occurring variations in wind and topography were indirectly included into the model predictions. Thus, this ability to fly in the migratory direction (the rstep-value) reflects a sum of uncertainties originating from both the orientational system itself and from wind and topographical effects. The length of the sample mean vector after one migratory step was found to be 0.665, equivalent to an angular standard deviation of ± 52° . This rstep-value was then inserted into the mathematical model and a theoretical curve emerged predicting where the ringing recoveries should be found (in terms of angular concentration around the mean migratory direction) as a function of distance migrated (see Fig. 3).
Then, all recoveries of Pied Flycatchers ringed in Scandinavia (Finland, Norway, Sweden and Denmark) until 1996 were analysed (for examples see Fig. 4). Only birds ringed and recovered within the same autumn north of the Sahara (n = 1138) were used in the analysis. The directional distributions were calculated (sample mean vector lengths, r-values) in 36 different distance intervals and plotted on the same figure (Fig. 3) as the model predictions for direct comparison.
From Fig. 3 it is evident that the directional concentrations (r-values) found in ringing recoveries and the equivalent predictions of the model correlate strongly. Thus, a simple clock-and-compass model is sufficient to describe the migratory pattern of Pied Flycatchers. If true navigational abilities were necessary and used by night migrating passerines, the directional distributions derived from ringing recoveries would be more concentrated, at least beyond some distance, than those predicted from the simple clock-and-compass model. Consequently, if the birds navigated, the theoretical curve should lie significantly under the dots derived from ringing recoveries. The directional concentrations derived from ringing recoveries were not significantly higher than those predicted by the model (Wilcoxon’s signed rank test: T- = 238, n = 35, P > 0.20).
Another way to visualise the results is shown in Fig. 5. Here, all birds were imagined to start out at the large dot (all recovery sites were parallel displaced as if the birds were ringed at the large dot). Then, the model predicted 66% of the recoveries to be found within the parabola. The actual recoveries were found to fit the model predictions extremely well, since 65% were located within the parabola. In fact, this similarity is higher than should be expected even if the model is correct.
In conclusion, the modelling study showed that Pied Flycatchers equipped only with an internal clock and a compass but no map will show directional distributions as a function of distance almost identical to the distributions found in ringing recovery data.
Displacement experiments with young Pied Flycatchers
The strong correlation found in the modelling study suggested that young Pied Flycatchers do not need compensatory abilities. However, the result did not exclude the possibility that young Pied Flycatchers may be able to compensate, if they make very large mistakes or drift very far. (Still, the results strongly suggested that the vast majority of the Pied Flycatchers did not compensate during their first autumn migration. Thus, if compensatory abilities exist, then the vast majority of the birds did not make large enough mistakes or did not drift far enough to trigger such a system, thereby making it of limited biological significance).
Therefore, the aim of the displacement study (Mouritsen & Larsen 1998) was to test whether young Pied Flycatchers caught in Scandinavia show compensatory orientation on their first migration when displaced about the maximum biologically realistic distance away from the normal migratory route (based on indications from ringing recoveries).
The study was designed to maximise changes of direction in case of compensatory orientation and to control for local variations. This was done by symmetrically displacing birds from a population of south-west migrating Pied Flycatchers towards west and south and measure their orientation relative to a control group at the site of capture (Fig. 6). In the case of clock-and-compass behaviour the birds should indicate the same migration direction in all three places. In case of compensatory behaviour, however, the prediction would be that birds displaced to the west and to the south should show the same (large) numerical change of direction towards the normal migration route while birds kept at the site of capture should keep the south-west orientation.
First, a total of 48 young Pied Flycatchers were caught and tested for directional preferences on Christiansø, Denmark. They oriented in the expected south-south-westerly direction. Second, they were split into three equal groups, one group was displaced about 500 km due south, another about 400 km due west, while the third remained as control at Christiansø. Then, the orientation of the three groups was tested simultaneously by three different experimenters. The birds were found to show the same orientation at all localities (all 95% confidence intervals overlap, see Mouritsen & Larsen 1998), showing no signs of compensatory orientation (see Fig. 6 ).
These results supported the suggestions of the modelling study, and judged together they strongly suggested that young Pied Flycatchers - and perhaps night migrating passerines in general - are equipped with compasses only, but no map. If this suggestion holds, then all prerequisites (a compass and an internal clock) that young night migrating passerines need to orient during autumn migration seem to be presently known, at least at the behavioural level.
Seen in this light a review summarising and re-evaluating the results of previous experiments studying the directional component of the spatiotemporal orientation programmes of night migrating passerines seemed useful. Especially, since the reviews on spatiotemporal orientation programmes in the past (e.g. Berthold 1991; Gwinner 1996) have focused mainly on the time component, and because most if not all of these data cannot be used to distinguish between the clock-and-compass and navigation hypothesis, respectively.
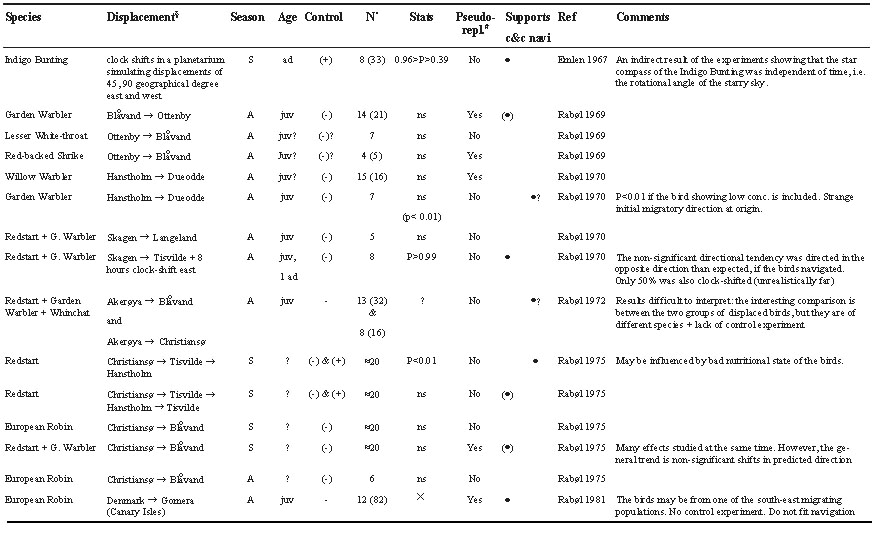
Reviewing and re-evaluating the literatureAll experiments investigating the compass component of the orientation programmes of night migrating passerines described in the literature were re-evaluated. The results are summarised in Table 1. For standardisation and comparative purposes, all statistical results were re-evaluated: The Watson-Williams test was used, if its underlying assumption was fulfilled: both sample mean vectors to be compared were ³ 0.75. If this was not the case, 95% confidence intervals were estimated using the charts in Batschelet (1981, page 86). The minimal sample size for Watson-Williams test is 6, whereas it is 8 for estimating confidence intervals. A given experiment was regarded as supporting the navigation hypothesis and a · was placed in the column indicating support of the navigation hypothesis, if the sample size was sufficiently high and if a significant directional shift in concordance with navigational/compensatory orientation was found (the level of significance was given in the ‘statistics’ column in Table 1). Non-significant directional shifts were regarded as supportive of the clock-and-compass hypothesis (since no change in direction is the null hypothesis). However, if a non-significant directional shift of minimum 10° (the approximate precision of the funnel method; Mouritsen & Larsen, unpublished manuscript) occurred in the direction expected for navigational or compensatory orientation, a (· ) was placed in the column indicating support of the clock-and-compass hypothesis and ‘ns’ was given in the ‘statistics’ column of Table 1. A · in the column ‘supports c&c’ (the clock-and-compass hypothesis) means that the directional preferences of the birds before and after displacement were not significantly different and directed in the normal migratory direction. Thereby, the results were clearly in accordance with the clock-and-compass hypothesis. If no control experiment was performed at all, statistical calculations cannot be performed (indicated by a X in the ‘statistics’ column of Table 1), else a P-value indicating the likelihood that the directional choices were NOT changed significantly as predicted by the navigation hypothesis was given. If sample sizes were too small and/or if the birds showed very scattered and/or unexplainable directions before or after displacement, no · was given in either the clock-and-compass nor in the navigation column.
The degree to which control experiments have been performed varies strongly in the previous studies. To indicate this, a column labelled ‘control’ is presented in Table 1. A ‘+’ indicates that the displaced birds were tested prior to and after displacement and a control group remaining at the site of capture was tested simultaneously. A ‘(+)’ indicates that the same birds were tested both before and after displacement. A ‘(-)’ indicates that birds caught simultaneously (within one week) with the displaced birds acted as control group and a ‘-’ indicates that no birds were tested prior to displacement, but some ‘normal migratory direction’ was assumed.
From Table 1 it is evident that Blåvand re-occurs either as the origin or destination in many displacement experiments, especially in those suggesting navigational responses. In this context, it is important to know that all locally caught and typically south-west-migrating species like Pied Flycatchers, Redstarts, Phoenicurus phoenicurus, and Garden Warblers orient south-east in funnels when tested at Blåvand (Rabøl 1969, 1975, 1995; Mouritsen 1998a). An equivalent south-easterly orientation was also observed during release experiments (Mouritsen 1998a). There are at least four possible reasons for this orientation. First, this phenomenon may be caused by the birds being of Norwegian origin and by assuming such birds having a south-easterly directed migration leg in Scandinavia before turning south-west somewhere south of Denmark (see e.g. Rabøl 1969, 1995; Mouritsen 1998a). Second, it might be caused by some kind of local anomaly (for example some effect of the North Sea to the west and south-west of this test site). Third, most birds caught at Blåvand may have been surprised by bad weather over the sea to the west of the light house and their occurrences can be looked upon as emergency landings (see ‘emergency plan A’ below). This might be remembered and expressed in their later orientation as a reluctance towards turning west again. Finally, the south-east orientation could be interpreted as compensatory if the birds had recently been blown out of course by easterly winds.
In earlier displacement experiments the south-easterly orientation at Blåvand was interpreted as being compensatory resulting from displacement. However, the peculiar south-east orientation is also found in birds caught locally during high-pressure periods with almost no or changing winds and good weather (Mouritsen unpublished data), and it has been found to persist even after two weeks in captivity (Mouritsen 1998a). These findings speak in favour of some anomaly causing the unusual site specific south-east orientation in normally south-west migrating species caught and tested at Blåvand. Consequently, south-east orientation at Blåvand (regardless of whether Blåvand is the origin or destination of a displacement) does not qualify as valid evidence for the navigation hypothesis, since all locally caught birds (which have not been displaced) also show south-east orientation.
If we exclude all those data in which south-east orientation at Blåvand is crucial, we are left with only two studies providing evidence in favour of the navigation hypothesis (see Table 1): The first being the displacement of Garden Warblers (Rabøl 1970) from Hanstholm à Dueodde. However, the orientation prior to the displacement was atypical and the birds displaced were not tested at the origin. The second is the displacement of Redstarts from Christiansø à Hanstholm à Tisvilde (Rabøl 1975). In this study, the birds were transported for several days in succession and tested during the nights in between, leaving the birds little time to feed and rest. Therefore, the observed orientation could also be seen as an example of the reverse orientation often observed in lean birds (Sandberg et al. 1988; Sandberg 1994; Åkesson et al. 1996; Sandberg & Moore 1996). In both of the above mentioned studies (Rabøl 1970, 1975), other groups of birds provided evidence in favour of the clock-and-compass hypothesis.
CONCLUSIVE EXAMINATION
A thought-provoking quotation from one of the early papers on displacement experiments about interpreting data to be evidence for the navigation hypothesis seems to summarise the last couple of pages: ‘As Wallraff has emphasised, this would have very important consequences regarding the orientational mechanism. Therefore, one should not accept it before having excluded other possibilities as an explanation of the results’ (Rabøl 1972).
In conclusion, whether or not we exclude the Blåvand results, both theory, lab experiments, field data and evidence from the literature strongly support the notion that at least young Pied Flycatchers, and probably young night migrating passerines in general, do not navigate during their first autumn migration. Therefore, if the navigation hypothesis shall remain a likely alternative spatiotemporal orientation mechanism to the clock-and-compass hypothesis for young night migrating passerines, it needs to be supported by some very carefully designed experiments providing highly significant results. Future experiments should include large sample sizes of young birds of a single species only, provide clear pre-experimental predictions and pay meticulous attention to control groups. Also, Blåvand and probably all sites close to any coastline should be avoided as origins or destinations for displacements, since a general repulsive effect of coast lines even in funnel experiments may exist. Such a tendency may be suggested by experiments at Hanstholm, Tisvilde and Langeland (Rabøl 1970), but other experiments do not find such a repulsive tendency (e.g. Sandberg & Moore 1996).
Some previous displacement experiments have suggested that young night migrating passerines may possess navigational abilities (see Table 1). Therefore, it is my hope that the present review will not stop the discussion completely, but instead inspire researchers to perform further carefully designed displacement experiments. I believe that such experiments carried out by a number of people could bring more interesting and important knowledge about the spatiotemporal orientation programmes and reaction patterns of night migrating passerines.
IS THE CLOCK-AND-COMPASS STRATEGY SUFFICIENT?
The severe selection pressures on choosing the optimal migration route and wintering quarter (see e.g. Berthold et al. 1992) should have facilitated evolution of a more sophisticated orientation mechanism, if any possibility existed for such a system to develop (i.e. cues functioning like longitudes and latitudes). However, no reliable east-west navigational cue that birds can sense is known at present. The axes provided by the angle of magnetic inclination and the height above the horizon of the rotational point of the stars are both oriented north-south and are almost parallel to each other except at very high latitudes.
Is it really likely that a simple clock-and-compass mechanism is all there is to the story? No, probably not quite, but let us add two emergency plans to the simple clock-and-compass strategy.
Emergency plan A: In Western Europe, ‘If you find yourself over water at dawn with no land in sight in autumn, turn approximately 90 degrees towards the sunrise (this means from your present SW-heading to a SE-heading) and continue on this heading until you encounter land.’ The existence of such an emergency plan in autumn was strongly suggested by the radar observations of night migrating thrushes over the North Sea (Myres 1964). At dawn they turned their course from south-west to south-east if they were located over the sea far from land, whereas they continued their south-westerly orientation if they were located over land or close to the British coast. If one inspects a map of Europe (and Africa) it is easy to see that using ‘emergency plan A’ would be very adaptive especially to a south-west-migrant travelling from northern and central Europe through southern Spain to tropical Africa. Due to the predominant westerly winds in northern Europe, night migrating passerines may aim at a slightly more westerly course than the one that would bring them to their goal without these winds. Such a strategy will be safe and may be advantageous on average as long as the birds use ‘emergency plan A’. It is easily imaginable that selection has resulted in other versions of this emergency plan in other parts of the world. For instance, a turn towards northwest have been reported in bird species migrating down the American east coast (e.g. Able 1977). Similar emergency plans (turning towards or away from the sunrise) may also exist in spring.
Emergency plan B: ‘If the wind is too strong to implement ‘emergency plan A’, fly with the wind until you encounter land’ (e.g. Richardson 1991). The existence of this emergency plan or forced behaviour in the real world is clearly indicated by occurrences of rarities and vagrants. The best example probably is the North American passerines regularly turning up in Great Britain and Iceland during periods with predominantly westerly winds (Alström et al. 1991). Another example is the occurrence of common migrants like European Robins, Erithacus rubecula, and Blackcaps far away from their normal migratory route on locations like Shetland and the Faeroe Isles during periods with predominantly easterly winds (e.g. Rabøl 1985).
If the finding that young Pied Flycatchers do not navigate during migration and the existence of the emergency plans can be generalised to all night migrating passerines, then a coherent description of the first autumn migration of young night migrating passerines seems closer at hand: in autumn, these birds set out on their journey equipped with at least two genetically encoded compasses, a time schedule and a couple of emergency plans only. At present, we know that night migrating passerines have an inherent time schedule (for a review see Gwinner 1996) and that they possess at least two compasses (e.g. W. Wiltschko & Wiltschko 1991, 1996; Mouritsen 1998), a magnetic compass and a celestial compass. An elusive map sense and/or any presently unknown east-west navigational cue are not needed (see e.g. W. Wiltschko & Wiltschko 1996). In conclusion, at present the clock-and-compass hypothesis seems to be sufficient to account for all well-studied phenomena and clear-cut data on young night migrating passerines.
WHAT ARE THE ORIENTATIONAL STRATEGIES OF ADULT NIGHT MIGRATING PASSERINES?
In contrast to the extensive amount of data collected on young night migrating passerines on their first autumn migration, very little seems to be known regarding orientational strategies of birds on their first spring migration (the migration route is often different from the one followed in autumn; see e.g. Zink 1985; Alerstam 1990) and of older and experienced birds on both spring and autumn migration. In any case, it is of vital importance to understand in detail the basic inherited components used by naïve birds before looking at the orientation strategies used by adults. Although, the strategies described above for young birds are of course also available to experienced birds, learning almost certainly adds some degree of navigational abilities. Else, it is very hard to explain the high breeding, wintering and even stop-over site fidelity found in many species (see e.g. Cantos & Tellería 1994; Villard et al. 1995). Also, the common phenomenon known to all ringers banding birds regularly at the same location for several years that quite a number of birds are caught in almost the same net on the same date for several years in a row, speaks strongly in favour of learnt navigational abilities in experienced migrants.
In this regard, it is important to realise that night migrating passerines usually migrate at altitudes of 300 to 2000 m (Bruderer et al. 1995; Bruderer 1997) above ground-level. This means that they under optimal clear conditions can see a very wide1 (see footnote)area of land from which they have the potential of using familiar landmarks for navigation. This reduces the needed precision of the long-distance orientational system of experienced night migrating passerines, since they do not need to find a very small patch of land based on this system. Instead, they may switch to landmark navigation already up to 160 km from their final goal. If the landmark navigation is based primarily on features like conspicuous rocks and mountain ranges elevated above the general surface of the earth, the landmark navigation range can be increased considerably. If experienced migrants have capabilities to learn several fix-points using landmarks along the migration route(s), their orientational system may be one of a simple clock-and-compass mechanism with emergency plans combined with landmark navigation. Studies of food-caching species of birds have revealed impressive capabilities of remembering a large number of spatial locations and landmarks (e.g. Bednekoff & Balda 1997). The food-caching species are mainly residents (else they will have little benefit from their food-stores). It is therefore straightforward to suggest that migratory species possess similar capabilities, which are used for remembering the locations of good foraging and wintering sites on their migratory path.
ACKNOWLEDGMENTS
I wish to express my very special thanks to Jørgen Rabøl whose inspiration, interest, critique and scepticism have been a great help to me in preparing this paper. I am grateful to Ole Næsbye Larsen and Axel Michelsen for valuable comments on the manuscript. The Centre for Sound Communication is financed by the Danish National Research Foundation.
REFERENCES
Able, K.P. 1977. The orientation of passerine noctunal migrants following offshore drift. Auk 94: 320-330.
Able, K.P. & Cherry, J.D. 1986. Mechanisms of dusk orientation in White-throated Sparrows: clock-shift experiments. Journal of Comparative Physiology 159: 107-113.
Åkesson, S., Karlsson, L., Walinder, G. & Alerstam, T. 1996. Bimodal orientation and the occurrence of temporary reverse bird migration during autumn in south Scandinavia. Behavioural Ecology and Sociobiology 38: 293-302.
Alerstam, T. 1990. Bird Migration. Cambridge; Cambridge University Press: 420pp.
Alerstam, T. 1996. The geographical scale factor in orientation of migratory birds. The Journal of Experimental Biology 199: 9-19.
Alström, P., Colston, P. & Lewington, I. 1991. A field guide to the rare birds of Britain and Europe. London; HarperCollins: 448pp.
Batschelet, E. 1981. Circular Statistics in Biology. London; Academic Press: 370pp.
Beck, W. 1984. Die Zugorientierung des Trauerschnäppers (Ficedula hypoleuca Pallas). PhD Thesis, J.W. Goethe-Universität, Frankfurt a.M., Germany.
Bednekoff, P.A. & Balda, R.P. 1997. Clark’s Nutcracker spatial memory: many errors might not be due to forgetting. Animal Behaviour 54: 691-698.
Berthold, P. 1973. Relationships between migratory restlessness and migratory distance in six Sylvia species. Ibis 115: 594-599.
Berthold, P. 1975. Migration: Control and metabolic physiology. In: Farner, D.S. & King, J.R. (eds) Avian Biology. Vol.5; New York; Academic Press: 77-128.
Berthold, P. 1991. Spatiotemporal programmes and genetics of orientation. In: Berthold, P. (ed) Orientation in birds. Basel; Birkhäuser Verlag: 86-105.
Berthold, P. & Querner, U. 1988. Was Zugunruhe wirklich ist - eine quantitative Bestimmung mit Hilfe von Video-Aufnahmen bei infrarotlichtbeleuchtung. Journal für Ornithologie 129: 372-375.
Berthold, P., Helbig, A.J., Mohr, G. & Querner, U. 1992. Rapid microevolution of migratory behaviour in a wild bird species. Nature 360: 668-670.
Bruderer, B. 1997. The study of bird migration by radar. Part 2: Major achievements. Naturwissenschaften 84: 45-54.
Bruderer, B., Underhill, L. & Liechti, F. 1995. Altitude choice of night migrants in a desert area predicted by meteorological factors. Ibis 137: 44-55.
Cantos, F.J. & Tellería, J.L. 1994. Stopover site fidelity of four migrant warblers in the Iberian Peninsula. Journal of Avian Biology 25: 131-134.
Emlen, S.T. 1967. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part II. Mechanism of celestial orientation. Auk 84: 463-489.
Emlen, S.T. & Emlen, J.T. 1966. A technique for recording migratory orientation of captive birds. Auk 83: 361-367.
Gwinner, E. 1968. Artspezifische Muster der Zugunruhe bei Laubsängern und ihre mögliche Bedeutung für die Beendigung des Zuges im Winterquartier. Zeitschrift für Tierpsychologie 25: 843-853.
Gwinner, E. 1986. Circannual rhythms in the control of avian migrations. Advances in the Study of Behaviour 16: 191-228.
Gwinner, E. 1996. Circadian and circannual programmes in avian migration. The Journal of Experimental Biology 199: 39-48.
Gwinner, E. & Wiltschko, W. 1978. Endogenously controlled changes in migratory direction of the Garden Warbler, Sylvia borin. Journal of Comparative Physiology 125: 267-273.
Helbig, A.J. 1991. Dusk orientation of European Robins, Erithacus rubecula: the role of sun-related directional information. Animal Behaviour 41: 313-321.
Helbig, A.J. 1996. Genetic basis, mode of inheritance and evolutionary changes of migratory directions in palaearctic warblers (Aves: Sylviidae). The Journal of Experimental Biology 199: 49-55.
Helbig, A.J., Berthold, P. & Wiltschko, W. 1989. Migratory orientation of Blackcaps (Sylvia atricapilla): population-specific shifts of direction during the autumn. Ethology 82: 307-315.
Lohmann, K.J. & Lohmann, C.M.F. 1996. Orientation and open-sea navigation in sea turtles. The Journal of Experimental Biology 199: 73-81.
Moore, F.R. 1987. Sunset and the orientation behavior of migrating birds. Biological Reviews 62: 65-86.
Moreau, R.E. 1972. The Palaearctic-African bird migration system. London; Academic Press: 384pp.
Mouritsen, H. 1998a. Redstarts, Phoenicurus phoenicurus, can orient in a true-zero magnetic field. Animal Behaviour 55: 1311-1324.
Mouritsen, H. 1998b. Modelling migration: the clock-and-compass model can explain the distribution of ringing recoveries. Animal Behaviour 56: 899-907.
Mouritsen, H. & Larsen, O.N. 1998. Migrating young Pied Flycatchers, Ficedula hypoleuca, did not compensate for geographical displacements. The Journal of Experimental Biology 201: 2927-2934.
Munro, U., Wiltschko, W. & Ford, H.A. 1993. Changes in the migratory direction of Yellow-faced Honeyeaters, Lichenstomus chrysops, during autumn migration. Emu 93: 59-62.
Myres, M.T. 1964. Dawn ascent and reorientation of Scandinavian thrushes (Turdus ssp.) migrating at night over the northeastern Atlantic Ocean in autumn. Ibis 106: 7-51.
Perdeck, A.C. 1958. Two types of orientation in migrating Sturnus vulgaris and Fringilla coelebs as revealed by displacement experiments. Ardea 46: 1-37.
Rabøl, J. 1969. Orientation of autumn migrating Garden Warblers (Sylvia borin) after displacement from Western Denmark (Blåvand) to Eastern Sweden (Ottenby). A preliminary experiment. Dansk Ornitologisk Forenings Tidsskrift 63: 93-104.
Rabøl, J. 1970. Displacement and phaseshift experiments with night-migrating passerines. Ornis Scandinavica 1: 27-43.
Rabøl, J. 1972. Displacement experiments with night-migrating passerines (1970). Zeitschrift für Tierpsychologie 30: 14-25.
Rabøl, J. 1975. The orientation of night-migrating passerines without the directional influence of the starry sky and/or the earth magnetic field. Zeitschrift für Tierpsychologie 38: 251-266.
Rabøl, J. 1978. One-direction orientation versus goal area navigation in migratory birds. Oikos 30: 216-223.
Rabøl, J. 1979. Magnetic orientation in night-migrating passerines. Ornis Scandinavica 10: 69-75.
Rabøl, J. 1981. The orientation of Robins Erithacus rubecula after displacement from Denmark to Canary Islands, autumn 1978. Ornis Scandinavica 12: 89-98.
Rabøl, J. 1985. The orientation of vagrant passerines on the Faeroe Islands, September 1984. Dansk Ornitologisk Forenings Tidsskrift 79: 133-140.
Rabøl, J. 1992. Star navigation in night-migrating passerines. Dansk Ornitologisk Forenings Tidsskrift 86: 178-181.
Rabøl, J. 1993. The orientation system of long-distance passerine migrants displaced in autumn from Denmark to Kenya. Ornis Scandinavica 24: 183-196.
Rabøl, J. 1994. Compensatory orientation in Pied Flycatchers, Ficedula hypoleuca, following a geographical displacement. Dansk Ornitologisk Forenings Tidsskrift 88: 171-182.
Rabøl, J. 1995. Compensatory orientation in juvenile Garden Warblers, Sylvia borin, and Redstarts, Phoenicurus phoenicurus, following a geographical displacement. Dansk Ornitologisk Forenings Tidsskrift 89: 61-65.
Richardson, W.J. 1991. Wind and orientation of migrating birds: a review. In: Berthold, P. (ed) Orientation in birds. Basel; Birkhäuser Verlag: 226-249.
Sandberg, R. 1994. Interaction of body condition and magnetic orientation in autumn migrating Robins, Erithacus rubecula. Animal Behaviour 47: 679-686.
Sandberg, R. & Moore F.R. 1996. Migratory orientation of Red-eyed Vireos, Vireo olivaceus, in relation to energetic condition and ecological context. Behavioural Ecology and Sociobiology 39: 1-10.
Sandberg, R., Pettersson, J. & Alerstam, T. 1988. Why do migrating Robins, Erithacus rubecula, captured at two nearly stop-over sites orient differently? Animal Behaviour 36: 865-876.
Sauer, E.G.F. 1957. Die Sternenorientierung nächtlich ziehender Grasmücken (Sylvia atricapilla, borin und curruca). Zeitschrift für Tierpsychologie 14: 29-70.
Villard, M.-A., Merriam, G. & Maurer, B.A. 1995. Dynamics in subdivided populations of neotropical migratory birds in a fragmented temperate forest. Ecology 76: 27-40.
Wiltschko, R. & Wiltschko, W. 1978. Relative importance of stars and the magnetic field for the accuracy of orientation in night-migrating birds. Oikos 30: 195-206.
Wiltschko, W. & Wiltschko, R. 1991. Magnetic orientation and celestial cues in migratory orientation. In: Berthold, P. (ed) Orientation in birds. Basel; Birkhäuser Verlag: 16-37.
Wiltschko, W. & Wiltschko, R. 1996. Magnetic orientation in birds. The Journal of Experimental Biology 199: 29-38.
Zink, G. 1985. Der Zug europäischer Singvögel - ein Atlas der Wiederfunde beringter Vögel. 4. Lieferung; Möggingen; Vogelzug-Verlag.
1The distance to the horizon seen by a 2 meter tall human being on a sphere with the radius of the earth, rearth = 6378 km at equator, is: rearth * inv cos((rearth) (rearth + 0.002 km)-1) » 5 km due to the curvature of the earth. A bird migrating at 500 m and 2000 m above ground level will, given perfect visibility, be able to see land within circles with radius 80 km and 160 km, respectively.
Table 1. Summary of previous experiments with night migrating passerines testing the clock-and-compass hypothesis against the navigation/compensation hypothesis. Further explanations see ‘reviewing and re-evaluating the literature’.
Table 1. continued.
Fig. 1. A
graphical representation of the mathematical model. First, a direction is picked randomly
from a Wrapped Cauchy Distribution characterized by the sample mean vector length, rstep.
A vector of length 1 in the picked direction is saved. Independently of the direction of
the first vector, a second direction is picked at random from the same distribution. A
vector of length 1 in the second picked direction is added to the first vector. This
procedure is followed n times, where n denotes the number of migratory steps. Finally, the
sum-vector is projected on the x-axis which represents the mean direction to the goal. The
resulting sample mean vector after n independent migratory steps, rn, is found
by dividing the length of the projection of the sum-vector on the x-axis, ![]() , with
the length of the sum-vector,
, with
the length of the sum-vector, ![]() . The procedure is repeated 100000 times for each
combination of n and rstep. This is mathematically equivalent to storing all
the individual mean directions and then find the sample mean vector of these 100000
directions in the end, but the present procedure makes the program faster and requires
much less storage capacity. The mean of the rn-values results in a precise
(within ± 0.002) estimation of the expected r-value after n
independent migratory steps (each following a Wrapped Cauchy distribution described by rstep)
for birds using a clock-and-compass strategy. The average distance travelled after n
migratory steps is found as the average of |sum|. [ From
Mouritsen 1998b by permission of the publisher, Academic Press Limited]
.
. The procedure is repeated 100000 times for each
combination of n and rstep. This is mathematically equivalent to storing all
the individual mean directions and then find the sample mean vector of these 100000
directions in the end, but the present procedure makes the program faster and requires
much less storage capacity. The mean of the rn-values results in a precise
(within ± 0.002) estimation of the expected r-value after n
independent migratory steps (each following a Wrapped Cauchy distribution described by rstep)
for birds using a clock-and-compass strategy. The average distance travelled after n
migratory steps is found as the average of |sum|. [ From
Mouritsen 1998b by permission of the publisher, Academic Press Limited]
.
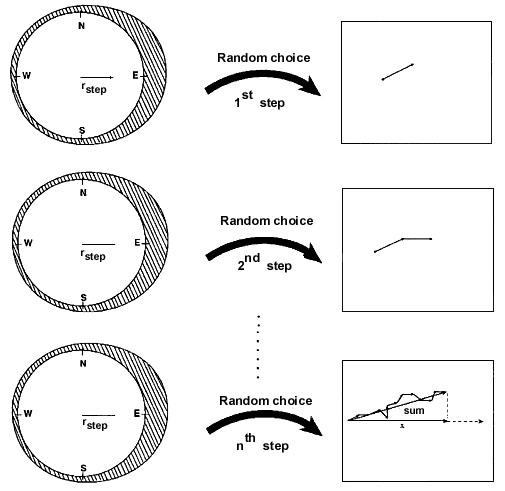
Fig. 2. The directional distributions of the birds recovered 100-149 km - equivalent to one migratory step - from the banding sites. Each dot represent one ringing recovery and indicate the direction from the banding to the recovery site. The sample mean vector is depicted as a line originating from the circle centre. ‘r’ indicates its length (‘r’ ranges between 0 and 1, being inversely related to the angular standard deviation). The inner and outer broken circles indicate the radii equal to significance levels of P = 0.05 and P = 0.01, respectively. Significance levels are according to the Rayleigh test (Batschelet 1981).
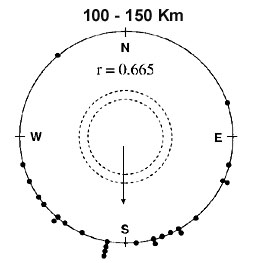
Fig. 3. Comparison between the results of the mathematical simulation of migration following the clock-and-compass model and data from natural migration obtained from recoveries of Pied Flycatchers ringed in Scandinavia within the same autumn. The bars show the number of recoveries in each distance interval. The error bars give the standard deviations [From Mouritsen 1998b by permission of the publisher, Academic Press Limited] .
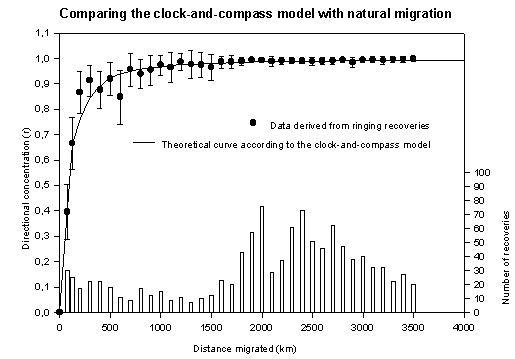
Fig. 4. Examples of the directional distributions of the ringing recoveries in eight of the 36 distance intervals. The other distributions are very similar to the ones shown. The lengths of sample mean vectors like these are the basis for the data points in Fig. 3. The distributions in eight other intervals can be found in Mouritsen (1998b). Symbols as described for Fig. 2.
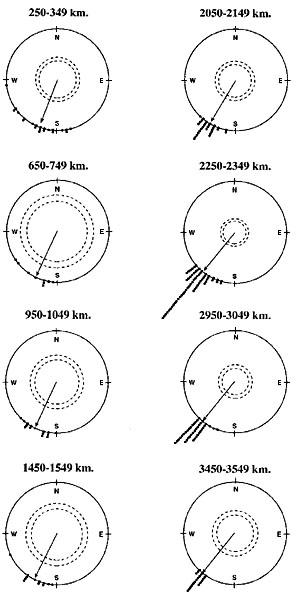
Fig. 5. Visualising the comparison between the predictions from the mathematical model of the simple clock-and-compass strategy and recoveries of Pied Flycatchers ringed in Scandinavia and recovered within the same autumn. The parabola shows the area within which the model predicts 66% (± 1 S.D.) of the ringing recoveries should be found, if young Pied Flycatchers use the simple clock-and-compass strategy and they all started at the location of the large dot (This location is approximately the centre of mass for the banding sites of the Pied Flycatchers included in the study of Mouritsen, 1998b). The small dots show where ringed birds were actually found (all banding and recovery sites were parallel displaced to have origin at the large dot) after 300, 500, 1000, 1500, 2000, 2500 and 3000 km. Just for visualisation purposes, the parabola and the recoveries (Fig. 4) are depicted upon a map of Europe. However, it is important to note that the birds, which seem to have been recovered over water, were in fact recovered on land. These birds were ringed somewhere away from the large dot. Their apparent recovery over water stem from imagining all birds to be banded at the location of the large dot, which resulted in an equivalent parallel displacement of all the recovery sites. The map is a Mercator projection of Europe.
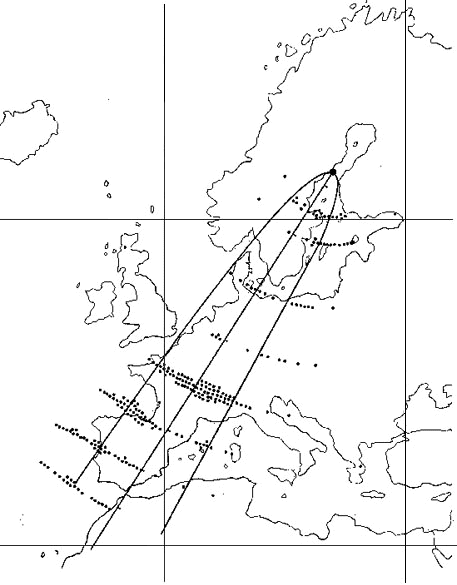
Fig. 6. Results of funnel (a-c, e-i) and release (d) experiments with first-year Pied Flycatchers during autumn migration before and after displacement from Christiansø, Denmark. The dots indicate the mean orientation of the migratory restlessness in funnels or vanishing bearings exhibited by the individuals that passed the exclusion rules. The sample mean vector is depicted as a line originating from the circle centre. The length of the sample mean vector is proportional to the angular concentration. The inner and outer dashed circles indicate the radii equal to significance levels of P=0.05 and P=0.01, respectively, according to the Rayleigh test (Batschelet 1981). The large bold arrows show the directions of the displacements (actual direction and distance are shown above the arrows). The bold dashed arrow indicates the normal migratory direction of Scandinavian Pied Flycatchers. [From Mouritsen & Larsen 1998, by permission of the publisher, The Company of Biologists Limited.]
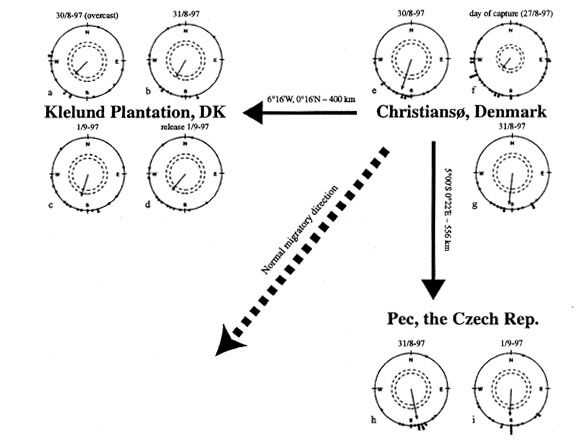
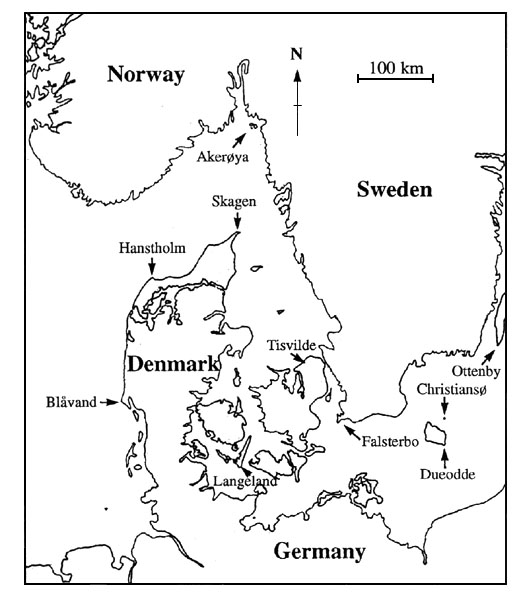
Fig. 7. Map showing the localities mentioned in Table 1 and in the text.