S16.2: Niche expansion and exploratory behaviour on islands: Are they linked?
Claudia Mettke-Hofmann
Forschungsstelle für Ornithologie der Max-Planck-Gesellschaft, Arbeitsbereich Biologische Rhythmen und Verhalten, Von der Tann Straße 7, 82346 Andechs, Germany, e-mail
Mettke@erl.ornithol.mpg.de Mettke-Hofmann, C. 1999 Niche expansion and exploratory behaviour on islands: Are they linked? In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 878-885. Johannesburg: BirdLife South Africa.Islands have a distinct biota which is characterised by an impoverished species community where some species are very abundant and where niche breadth often expands. There are several hypotheses which explain how increased population sizes could have occurred (competitive release, spill over, local adaptations). Niche expansion might be caused by specific island conditions such as less competition, low predation pressure, qualitative and quantitative differences in resources and other factors. However, little is known about how the behavioural processes permitting niche expansion. Exploratory behaviour might be one way in which species adapt to the new environment. As it will be shown island species are more explorative than mainland species. On islands animals are able to discover new resources or to develop new skills through exploration without a higher risk of predation but probably with a higher pay off than on the mainland because of there are fewer competitors. Thus, exploratory behaviour might be an important behavioural trait permitting niche expansion.
Islands differ greatly in their geographical range, their size, altitude, and other characters, but in general they have in common that they are more or less separated from greater landmasses and so can not easily be reached by terrestrial animals. As a result island communities differ in several aspects from nearby mainland communities.
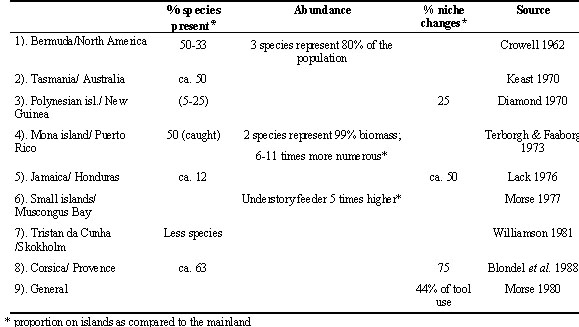
One phenomenon often observed on islands is a remarkably impoverished species diversity holding only around 50% of species compared to similar mainland areas (Table 1). Despite the low number of species on islands some species can be very abundant. Niche changes occur in 25 to 75 percent of the island species as compared to their mainland relatives (Table 1). These changes may consist of a niche shift, where the whole niche is shifted in one or the other direction so that the mean value of a niche use changes, or of a niche enlargement or narrowing, where the mean value remains the same but the variance increases or decreases (Williamson 1981). Niche changes can concern a variety of environmental conditions. Island species expand into new habitats, use a broader range of altitudes, increase their feeding spectrum, use a wider range of vertical feeding strata, or change their behaviour (Table 2). Several species developed new skills mainly linked to the context of feeding. For example, various island rails from different genera and the Galapagos Mockingbird Nesomimus sp. developed skills to break eggs of other birds (Amadon 1966), the Sharp-beaked-Groundfinch Geospiza difficilis feed on blood by pecking the secondaries of incubating bobbies Sula sp. (Bowman & Billeb 1965). Additionally, tool use behaviour evolved in 44% of all known cases on islands or island like areas (Morse 1980). A well known example is the Woodpecker finch Camarhynchus pallidus, which uses a spine to dislodge prey from crevices (Eibl-Eibesfeldt 1961).
What causes these changes and how can they be achieved?
Let us first consider some conditions on islands which may be different from the mainland.
As mentioned earlier, islands are often species impoverished which may be due to limited dispersal abilities of the species (Mac Arthur & Wilson 1963) or to ecologically less differentiated island habitats (Lack 1976). Thus, island species experience less interspecific competition (competitive release, Diamond 1970; Keast 1970; Mac Arthur et al. 1972; Terborgh & Faaborg 1973; Yeaton & Cody 1974; Morse 1977; Baker-Gabb 1986; Keeler-Wolf 1986; Löfgren 1995), which opens the possibility to occupy vacant niches.
Species impoverishment not only influences species interactions at the same trophical level but between different trophical levels as well. Typically, islands carry less predators than the mainland or they are predator free, especially from ground predators (Morse 1971; McLaughlin & Roughgarden 1989; Keast 1991; Daugherty et al. 1993; Blazquez et al. 1997). Flightlessness in birds and insects is a well known phenomenon on islands (Grant 1998) and on many islands animals are less cautious.
Food resources often differ in quality and quantity on islands from those on nearby mainland (Diamond 1975; Stamps & Buechner 1985). For example, insectivorous animals on islands at a lake in Sweden have to cope with a smaller abundance of arthropods than on the surrounding mainland. Additionally, the decrease in abundance is unequally distributed among different size classes (Nilsson & Ebenman 1981). Low abundance in resources facilitates a broader feeding range (Cody 1974; Pulliam 1986). But arthropod abundance of some specific guilds can also be increased as is the case for the foliage arthropods on Tobago island (Keeler-Wolf 1986). Further, seasonality and predictability of resources are important factors in determining niche shape (Cody 1974). Milder climatic conditions, especially on mediterranean islands, provide a more constant and predictable food supply as compared to the mainland (Morse 1977; Blondel et al. 1988) thus facilitating the specialisation on specific food sources. Many investigations show that despite the fact of a broader feeding niche of a particular species on islands, each individual is more specialised than on the mainland (Ebenman & Nilsson 1982; Werner & Sherry 1987). Additionally, the higher population densities often observed on islands are put down to a more stable food supply (Crowell 1962; Mac Arthur et al. 1972; Blondel et al. 1988).
Of course, there are many more factors in which islands differ from the mainland and on each island different constellations of factors will work. Additionally, species react differently to these changes as I will show in the following.
Some species become extremely abundant on islands and show niche expansions. Obviously, these species found optimal conditions on their particular islands. The increase in abundance may be due to higher resource availability because of reduced interspecific competition for the same resources (Crowell 1962; Stamps & Buechner 1985), or to better climatic conditions (Blondel et al. 1988). Several hypotheses exist as to why habitat expansion occurred in these species. One possibility is that suboptimal habitats could be inhabitat because of the lack of competitors (competitive release). For example, on Mona island Catbirds Dumetella carolinensis, Cardinals Richmondena cardinalis, and White-eyed Vireos Vireo griseus expanded their breeding range into more open habitats (Crowell 1962). As Crowell pointed out less competition facilitated these expansions.
Another possibility is that the population size of these species increased first in the optimal habitat. Because of the island situation, increasing intraspecific competition and restricted dispersal (fence effect, Krebs et al. 1969) lead to a 'spill over' into suboptimal habitats (spill over hypothesis, Crowell 1983; Blondel et al. 1988). This is the case in the Corsican island (Blondel et al. 1988) where some forest species show heavily increased population densities in the forest and a marked habitat expansion into matorral scrubland habitats.
A third explanation is given by Martin (1992) who reanalysed the data from Corsica from an autecologically point of view. Niche expansion might be possible because of local adaptations to the environmental conditions under geographical isolation. Therefore, the Corsican forest birds adjusted their breeding biology to the later onset of spring on the island. This delay in breeding facilitated them to be successful in matorral habitat as well.
Other species may not find such optimal conditions on islands. They do not increase in abundance but niche expansions occur in different dimensions caused by several reasons. In many of these cases reduced interspecific competition are thought to be the reason for niche expansion. On several small islands north east of New Guinea species expanded into higher altitudes normally occupied by other species (Diamond 1970). Another reason why niche expansion occurred in these species is that the habitat types differ between island and mainland. Wrens on St. Kilda and Fair Isle Troglodytes t. hirtensis and fridariensis show a remarkable shift in breeding habitat from vegetated areas on the mainland to treeless rocky sea cliffs on the islands (Williamson 1981). Resource scarcity or changed resources may be another factor causing niche expansion. Examples are the expansion of feeding niches in Galapagos birds which were sometimes accompanied by new feeding techniques as already mentioned.
For completeness there are also species showing no changes on islands at all, whereas others decrease their niche in one dimension, for example the habitat type, because they can use a wider range of resources exploited by other species on the mainland (Blondel et al. 1988).
Which are the behavioural processes permitting niche expansion?
First, habitats, food resources, or behavioural patterns already but infrequently used on the mainland because of the presence of other species, can become regularly used on islands in the absence of these competitors, thus leading to a broadening of the niche. In Willow Warblers Phylloscopus trochilus an island population in southern Sweden feeds more frequently at or near the trunk without changes in occurrence and frequency in feeding techniques as compared to the mainland population (Nilsson & Ebenman 1981).
Second, new resources or behavioural patterns can be achieved by observational learning. Werner & Sherry (1987) stated that individual feeding specialisation in Cocos finches Pinaraloxias inornata may be achieved by observational learning from other group members or even other species. As a whole this species show an extremely broad food spectrum.
Third, exploratory behaviour can play an important role in acquiring broader niches. New habitats or resources can be discovered through exploration. Do island species exhibit pronounced exploratory behaviour?
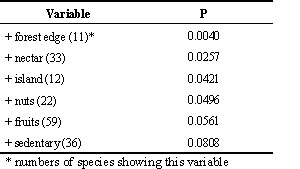
There are only few investigations concerning exploratory behaviour of island and mainland species. Haemig (1989) found that Scrub jays Aphelocoma coerulescens on Santa Cruz island approached a noise caused by rubbing on a ball more closely and stayed for a longer time at the source of noise than their Californian relatives. Thus, the island birds seem to be more explorative than the mainland birds. A second investigation was carried out with parrots. In this study, exploratory behaviour of 61 parrot species Psittacidae was investigated in captivity and correlated to their ecological requirements in the wild with respect to habitat use, food spectrum, origin from island or mainland, and other factors (Mettke 1995). Among others the variable 'island-mainland' showed an influence on exploratory behaviour (Table 3). Island species investigated a new object, a wooden ring, in their familiar aviary earlier than mainland species. Thus, the former reacted very quickly to changes in their environment which enable them to adapt their behaviour to new situations early in time. Again, island species were more explorative than mainland species.
There are several factors on islands which favour exploratory behaviour. First, predation pressure is reduced on islands. To be more explorative is therefore not correlated with increased costs of predation. Second, temporary food shortages can occur on islands. Discovering new resources in time through exploration helps to survive such bottlenecks (Haemig 1989). Third, less competing species increase the chance that newly discovered resources can successfully be exploited. And fourth, as already mentioned, resources on islands are often different from those on the mainland. Explorative species are able to find new resources earlier than less explorative ones. This might be important in island colonisation. Thus, the specific island conditions favour exploratory behaviour.
Is exploratory behaviour on islands linked with niche expansion on islands, in other words, do species with a strong niche expansion show more exploratory behaviour? The answer is still unknown. But in the genus Chalcopsitta (Psittacidae) the mainland species’ show a smaller food niche than the island species (C. cardinalis) and the former are less explorative than the latter. Also, the more curious Santa Cruz island Scrub jay investigated by Haemig (1989) occupies a broader range of habitats than their mainland relatives (Atwood 1980). Additionally, van Lawick-Goodall (1976) and Westergaard (1992) showed in chimpanzees and baboons that object exploration can lead to later tool-use behaviour which evolved more often on islands than on the mainland (Morse 1980).
For the future it will be interesting to focus more on the mechanisms of niche change. The investigation of exploratory behaviour may shed some light upon why niche expansion occurs in some species but not in others.
ACKNOWLEDGEMENTS
I would like to thank Prof. Dr. H. Winkler for financial support.
REFERENCES
Amadon, D. 1966 Insular adaptive radiation among birds. In: Bowman, I.R. (ed.) The Galapagos. Proceedings of the Symposia of the Galapagos International Scientific Project. University California Press: 18-30.
Atwood, J.L. 1980 Social interactions in the Santa Cruz island Scrub Jay. Condor 82: 440-448.
Baker-Gabb, D.J. 1986 Ecological release and behavioural and ecological flexibility in Marsh harriers on islands. Emu 86: 71-81.
Blazquez, M.C., Rodriguez-Estrella, R. & Delibes, M. 1997 Escape behaviour and predation risk of mainland and island Spiny-tailed Iguanas (Ctenosaura hemilopha). Ethology 103: 990-998.
Blondel, J., Chessel, D. & Frochot, B. 1988 Bird species impoverishment, niche expansion, and density inflation in mediterranean island habitats. Ecology 69: 1899-1917.
Bowman, R.I. & Billeb, S.L. 1965 Blood-eating in a Galapagos finch. The Living Bird: 29-44.
Carrascal, L.M., Moreno, E. & Valido, A. 1994 Morphological evolution and changes in foraging behaviour of island and mainland populations of Blue Tit (Parus caeruleus) – a test of convergence and ecomorphological hypotheses. Evolutionary Ecology 8: 25-35.
Cody, M.L. 1974 Competition and the structure of bird communities. Monographs in Population Biology 7.
Crowell, K.L. 1962 Reduced interspecific competition among the birds of Bermuda. Ecologia 43: 75-88.
Crowell, K.L. 1983 Islands – insight or artifact?: Population dynamics and habitat utilization in insular rodents. Oikos 41: 442-454.
Daugherty, C.H., Gibbs, G.W. & Hitchmough, R.A. 1993 Mega-island or micro-continent? New Zealand and ist fauna. Trends in Ecology and Evolution 8: 437-442.
Diamond, J.M. 1970 Ecological consequences of island colonization by southwest pacific birds, I. Types of niche shifts. Proceedings of the National Academy of Science 67: 529-536.
Diamond, J.M. 1975 Assembly of species communities. In: Cody, M.L. & Diamond, J.M. (eds) Ecology and evolution of communities. Cambridge, Massachusetts, London; Belknap Press of Harvard University Press: 342-444.
Ebenman, B & Nilsson, S.G. 1982 Components of niche width in a territorial bird species: Habitat utilization in males and females of the Chaffinch (Fringilla coelebs) on islands and mainland. The American Naturalist 119: 331-344.
Eibl-Eibesfeldt, I. 1961 Ueber den Werkzeuggebrauch des Spechtfinken (Camarhynchus pallidus). Zeitschrift fuer Tierpsychologie 18: 343-346.
Grant, P.R. 1998 Patterns on islands and microevolution. In: Grant, P.R. (ed) Evolution on islands. Oxford, New York, Tokyo; Oxford University Press: 1-17.
Haemig, P.D. 1989 A comparative experimental study of exploratory behaviour in Santa Cruz island and mainland California Scrub Jays Aphelocoma coerulescens. Bird Behaviour 8: 38-42.
Keast, A. 1970 Adaptive evolution and shifts in niche occupation in island birds. Biotropica 2: 61-75
Keast, A. 1991 Avian evolution of southern pacific island groups: An ecological perspective. Acta XX Congressus internationalis ornithologici 1990 Christchurch, New Zealand: 435-446.
Keeler-Wolf, T. 1986 The Barred Antshrike (Thamnophilus doliatus) on Trinidad and Tobago: Habitat niche expansion of a generalist forager. Oecologia 70: 309-317.
Kingdon, J. 1990 Island Africa: The evolution of Africa’s rare animals and plants. London, Glasgow, Sydney, Auckland, Toronto, Johannesburg; Collins Sons & Co Ltd..
Krebs, C.J., Keller, B.L. & Tamarin, R.H. 1969 Microtus population biology: Demographic changes in fluctuating populations of M. ochrogaster and M. pennsylvanicus in southern Indiana. Ecology 50: 587-607.
Lack, D. 1976 Island biology illustrated by the land birds of Jamaika. Oxford, London, Edinburgh, Melbourne; Blackwell Scientific Publications.
Lawick-Goodall, J. van 1976 Early tool using in wild Chimpanzees. In: Bruner, J.S., Jolly, A & Sylva K. (eds) Play – Its role in development and evolution. Harmondworth Middlesex; Penguin books: 222-225.
Löfgren, O. 1995 Niche expansion and increased maturation rate of Clethrionomys glareolus in the absence of competitors. Journal of Mammalogy 76: 1100-1112.
MacArthur, R.H. & Wilson, E.O. 1963 An equilibrium theory of insular zoogeography. Evolution 17: 373-387.
MacArthur, R.H., Diamond, J.M. & Karr, J.R. 1972 Density compensation in island faunas. Ecology 53: 330-342.
Martin, J.-L. 1992 Niche expansion in an insular bird community: an autecological perspective. Journal of Biogeography 19: 375-381.
McLaughlin, J.F. & Roughgarden, J. 1989 Avian predation on Anolis lizards in the northeastern Caribbean: An inter-island contrast. Ecology 70: 617-628.
Mettke, C. 1995 Explorationsverhalten von Papageien – Adaptation an die Umwelt? Journal fuer Ornithologie 136: 468-471.
Morse, D.H. 1971 The foraging of warblers isolated on small islands. Ecology 52: 216-228.
Morse, D.H. 1977 The occupation of small islands by passerine birds. The Condor 79: 399-412.
Morse, D.H. 1980 Behavioural mechanisms in ecology.Cambridge, Massachusetts, London; Harvard University Press.
Nilsson, S.G. & Ebenman, B. 1981 Density changes and niche differences in island and mainland Willow warblers Phylloscopus trochilus at a lake in southern Sweden. Ornis Scandinavica 12: 62-67.
Pulliam, H.R. 1986 Niche expansion and contraction in a variable environment. American Zoologist 26: 71-79.
Sheppard, D.H., Klopfer, P.H. & Oelke, H. 1968 Habitat selection: Differences in stereotypy between insular and continental birds. The Wilson Bulletin 80: 452-457.
Stamps, J.A. & Buechner, M. 1985 The territorial defence hypothesis and the ecology of insular vertebrates. The Quarterly Review of Biology 60: 155-181.
Terborgh, J. & Faaborg, J. 1973 Turnover and ecological release in the avifauna of Mona island, Puerto Rico. The Auk 90: 759-779.
Werner, T.K. & Sherry, T.W. 1987 Behavioural feeding specialization in Pinaroloxias inornata, the 'Darwin’s Finch' of Cocos island, Cosa Rica. Proceedings of the National Academy of Science 84: 5506-5510.
Westergaard, G.C. 1992 Object manipulation and the use of tools by infant baboons (Papio cynocephalus anubis). Journal of Comparative Psychology 106: 398-403.
Williamson, M. 1981 Island populations. Oxford; Oxford University Press.
Yeaton, R.I. & Cody, M.L. 1974 Competitive release in island Song sparrow populations. Theoretical Population Biology 5: 42-58.
Table 1. Overview over some typical differences between mainland and island communities
Table 2. Types of niche changes on islands.

Table 3. Influence of ecological variables on exploration latencies of 61 different parrot species. The table shows which ecological variables out of 13 influence exploration latencies (multiple regression analysis, n=61 species, r²= 0.348).