S15.3: Behavioural ecology of migratory orientation
Roland Sandberg & Susanne Åkesson
Department of Animal Ecology, Lund University, Ecology Building, S-223 62 Lund, Sweden, fax 46 46 2224716, e-mail Roland.Sandberg@zooekol.lu.se & Susanne.Akesson@zooekol.lu.se
Sandberg, R. & Åkesson, S. 1999. Behavioural ecology of migratory orientation. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 819-833. Johannesburg: BirdLife South Africa.Birds face several different behavioural challenges while on migration. Migrants must satisfy energetic demands of flight, which are especially high for species that have to cross ecological barriers like seas and deserts. Furthermore, birds have to correctly assess and integrate available directional information from environmental sources including the sun, the stars and the geomagnetic field. What factors influence the decision to migrate, choice of stopover sites and the orientation of migratory movements? Several recent studies, using both orientation cage and free-flight release techniques, have shown that: (1) The amount of stored fat has a major effect on the decision to migrate, lean birds being significantly less likely to embark on migration. (2) Fat stores can be used to predict whether an individual will select a seasonally appropriate direction or engage in reoriented movements. (3) Limited or nonexistent access to celestial orientation cues often leads to orientation in seasonally inappropriate directions. (4) Experienced long-distance migrants are significantly less scattered in their directional choices than are inexperienced conspecifics. (5) Migratory habits (inter- vs. intra-continental migration) may influence orientational precision. (6) Conspecifics in different migratory situations with respect to ecological barriers assign different priorities to visual and nonvisual orientation cues. The results emphasise the need to take into account ecological factors when analysing the complex orientation behaviour of migrating birds.
INTRODUCTION
Recent investigations concerning the behavioural basis of orientation systems of migratory birds have revealed a complicated web of interactions with physiological status and other ecological factors. Although migration allows individual birds to take advantage of different habitats as environments change seasonally (e.g. Alerstam and Enckell 1979; Alerstam and Högstedt 1982; Levey and Stiles 1992), it also involves several behavioural challenges that have to be met in a timely manner to ensure survival (Alerstam and Lindström 1990). Migrants must deal with potential costs including (a) high-level energetic demands of flight (Blem 1980; Pennycuick 1989; Norberg 1990), (b) foraging in unfamiliar habitats which may lead to increased exposure to predation (Lindström 1990; Moore 1994), (c) competition with other individuals at stopover sites which may result in impaired food acquisition (Hansson and Pettersson 1989; Moore and Yong 1991), (d) weather variations that influence energetic costs of flight, the maintenance of preferred headings and mortality en route (Richardson 1978, 1990; Alerstam 1981) and (e) the risk of orientation errors (Alerstam 1988, 1990).
In this paper, we consider the available experimental evidence concerning the influence of several ecological factors on the migratory orientation behaviour of birds. In doing so, we will attempt to answer the following questions: (1) How do the energetic stores of migrants influence the decision to embark on migration and the subsequent choice of a migratory direction? (2) How does the accessibility of different orientation cues affect directional choices (e.g. under clear skies and complete overcast)? (3) Do orientation behaviour and directional accuracy differ between experienced (old) and inexperienced (young) migrants? (4) How do migratory habits (i.e. long-distance vs. short-distance migrants) affect the orientation behaviour of migrants? (5) How is the integration of different directional information tied to the ecological context (i.e. migratory situation)?
INFLUENCE OF ECOLOGICAL FACTORS ON MIGRATORY ORIENTATION BEHAVIOUR
Energetic stores and the motivation to migrate
One of the most important requirements during migration is to meet energetic demands, particularly in relation to the crossing of ecological barriers (Wood 1982; Bairlein 1985; Biebach 1985, 1990; Biebach et al. 1986; Moore and Kerlinger 1987; Safriel and Lavee 1988; Dolnik 1990; Kuenzi et al. 1991). Migratory birds, especially when facing a barrier crossing, are expected to show well-oriented activity in the seasonally appropriate direction only if they have deposited adequate fat stores for long-distance flights. Individuals carrying insufficient energy stores should either suppress migratory activity if stopover conditions are benign (i.e. stay on their current location, cf. Moore and Kerlinger 1991; Sandberg et al. 1991; Yong and Moore 1993; Sandberg and Moore 1996) or, if feeding opportunities are restricted, reorient in search of more profitable feeding areas (Alerstam 1978; Terrill and Ohmart 1984; Lindström and Alerstam 1986; Sandberg et al. 1988a, 1991; Sandberg 1994; Åkesson et al. 1996; Sandberg and Moore 1996).
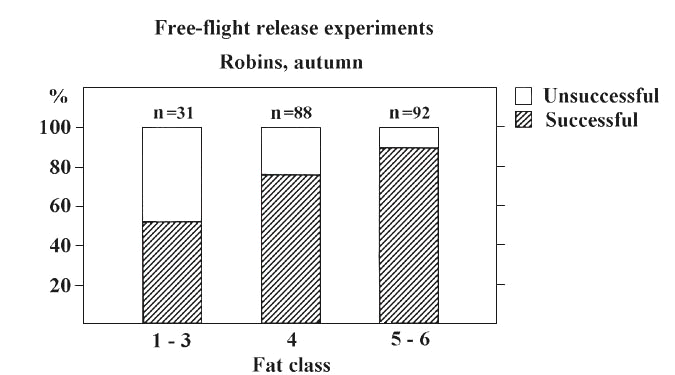
Recently, clear-cut evidence was presented by Sandberg et al. (1991) who performed free-flight release experiments where Robins Erithacus rubecula were released singly 1-2 h after local sunset. The birds which were carrying a small chemiluminescent lightstick were tossed into the air much in the same way as homing pigeons. Released birds were then followed by using 10x40 binoculars until they vanished from sight (for further details on methods, see Sandberg et al. 1991). The results show that the amount of stored fat has a strong effect on the birds' decision to take off on migratory flights or not (Fig. 1). Robins with high fat levels were significantly more likely to embark on autumn migration flights than were individuals with low fuel stores. This result has been confirmed in similar release experiments with Red-Eyed Vireos Vireo olivaceus where 38% of the lean birds decided to stay on their current location whereas 100% of the fat birds took off on migratory flights in the seasonally appropriate direction (Sandberg & Moore 1996). Furthermore, in those few cases when lean birds actually flew away from the release site, they most often reoriented in directions roughly opposite to the seasonally appropriate (Sandberg et al. 1991, Sandberg and Moore 1996). Finally, in orientation cage tests with Red-eyed vireos, fat individuals were significantly more inclined to display migratory activity (81%) than were lean birds (61%; Sandberg & Moore 1996).
Fuel stores and the choice of migratory direction
The influence of varying amounts of energetic stores on the migratory orientation of birds has received surprisingly little experimental attention. Able (1977) performed a study on a number of different North American warblers, using Emlen-funnels (cf. Emlen and Emlen 1966), and showed that the amount of visually estimated fat stores did not affect the quantity of migratory activity in orientation cages, but had a pronounced effect on the concentration of directional selections (i.e. fat birds were well oriented whereas lean individuals were not). Reoriented autumn migration, more or less in the opposite direction relative to the seasonally appropriate, is not uncommon when landbirds encounter an ecological barrier (e.g. Able 1977; Alerstam 1978; Richardson 1978; Bruderer and Jenni 1988; Sandberg et al. 1988a; Åkesson et al. 1996). Such a behaviour may constitute an adaptive response which permits migrants to search from profitable stopover sites away from densely populated coastal areas (avoidance of competitors and predators; cf. Alerstam 1978).
Recent experimental evidence for a clear connection between the amount of stored fat and directional selections stem from orientation cage studies using Robins (Sandberg et al. 1988a; cf. also Sandberg 1994), Chaffinches Fringilla coelebs (Bäckman et al. 1997), Red-eyed Vireos Vireo olivaceus (Sandberg and Moore 1996) and Snow Buntings Plectrophenax nivalis (Sandberg et al. 1998). For example, Snow Buntings tested in high arctic Canada (Resolute; Cornwallis Island) during the autumn migration period showed distinct differences in preferred mean orientation between fat and lean birds (Fig. 2; Sandberg et al. 1998). Similar clear-cut evidence has been obtained in free-flight release experiments with Pied Flycatchers Ficedula hypoleuca (Sandberg et al. 1991) and Red-eyed Vireos (Sandberg and Moore 1996).
Availability of different orientation cues affect directional choices
Radar and ceilometer studies have demonstrated that fewer birds embark on migration under solid overcast as compared to clear skies (e.g. Richardson 1978). Furthermore, with access to sunset and/or stellar information, birds usually orient in the expected direction for the season, even into headwinds, whereas migrants often seem to have difficulties in selecting a seasonally appropriate direction under solid overcast, and instead opt for downwind headings (Able 1978, 1980, 1982a,b).
Several experimental studies, including free-flight release experiments and orientation cage studies, have confirmed the importance of access to visual celestial cues for seasonally appropriate and well-oriented behaviour (e.g. Emlen and Demong 1978; Able et al. 1982; Sandberg 1991; Sandberg et al. 1991; Åkesson 1993,1994). For example, Robins released under clear autumn skies in South Scandinavia managed to select a well-defined mean direction towards the expected SW direction, in good agreement with available ringing data (Fig. 3, left diagram; Sandberg et al. 1991). In contrast, autumn releases under complete overcast resulted in strikingly different headings (Fig. 3, right diagram) with a significant cluster of vanishing bearings in the downwind direction. Moreover, the Robins required significantly longer time to select their vanishing directions in the absence of visual cue information. Studies performed by using orientation cages covered with white diffusing Plexiglass to simulate complete overcast conditions commonly result in random orientation and/or high angular dispersion of directional choices (Åkesson 1993, 1994), but not consistently so (cf. Sandberg and Pettersson 1996). Interestingly, visual information low over the horizon during the twilight period seems to be important for seasonally correct orientation. Robins tested in orientation cages with access to either 90° or 160° field of vision, centered around zenith, displayed significantly different orientation behaviour. When tested with a restricted field of vision (90° ) the Robins chose headings almost directly towards the position of the setting sun whereas individuals with more unrestricted visual access to the lower parts of the twilight sky oriented in their expected autumn migration direction (Sandberg 1991).
Influence of experience on migratory orientation
Yearling migrants are presumed to fly along inherited directions for an endogenously controlled time period (the so-called 'clock and compass hypothesis') to reach their population-specific wintering grounds during the first autumn migration, whereas second-year birds and adults in all seasons may use some form of bi-coordinate navigation to reach their migratory destinations (reviewed by: Berthold 1988, 1991; but see Rabøl 1985). Innate programs, of course, do not preclude an importance of experience. Conditions encountered en route and experiences during previous migratory journeys may modify orientation behaviour and play a role in the regulation of bird migration (Ketterson and Nolan 1985, 1988; Gauthreaux 1982; Terrill 1988). Migratory experience may affect the integration of different orientation cues in the determination of a direction (Keeton 1980; Sandberg 1990; Wiltschko 1991) as well as the maintenance of a preferred heading (Moore 1984; Able and Bingman 1987; Alerstam 1990; Sandberg et al. 1991).
Reports on orientation errors in the literature almost exclusively refer to migratory naive (inexperienced) birds on their first autumn migration (Drury and Keith 1962; Able 1977; Ralph 1978; DeSante 1983). This is further stressed by the fact that 'vagrancy' is most often connected with young birds found in coastal areas, far off their normal migration route (Alerstam 1990). Orientation errors could result from either an inability to follow a consistent compass course within a migratory season (disorientation), or from a failure to select the correct species-specific migratory direction (misorientation) (DeSante 1983; Alerstam 1990). Observed variability in the compass orientation of a population is a composite function of variation within and between individuals (Moore 1985). Individuals that consistently select seasonally appropriate headings would be at a selective advantage, especially in the light of the possibly dire consequences associated with orientational errors (Ralph 1978, 1981; Alerstam 1988, 1990).
Yearling migrants are more likely to become misoriented than adult birds (DeSante 1983; Ralph 1978; Williams and Williams 1978), and directional selections of juveniles often show greater angular dispersion than those of adults (e.g. Rabøl 1978; Moore 1984; Fransson 1986; Hedenström and Pettersson 1987; Sandberg 1990). For example, Sandberg et al. (1988a) reported that juvenile autumn migrating Robins showed a significantly larger scatter of headings than adults when tested in orientation cages at twilight. Similarly, in both Robins and Pied Flycatchers, adults displayed more concentrated vanishing directions than yearling birds during free-flight release experiments in southern Scandinavia (Sandberg et al. 1991). In Fig. 4, we have compiled ringing recovery data for Pied Flycatchers and Reed Warblers Acrocephalus scirpaceus, two intercontinental migrants, both of which show significantly greater angular dispersion of migratory directions for first-year birds as compared to adults.
Effects of migratory habits on orientation behaviour
Are there any differences in the concentration of preferred headings between intercontinental (long-distance) and intracontinental (short-distance) migrants? This question has received little, if any, attention in the scientific literature. The ample availability of ringing data, especially on Palaearctic migrants, makes it possible to investigate whether there are any differences in the orientation behaviour between long-distance and short-distance migrants.
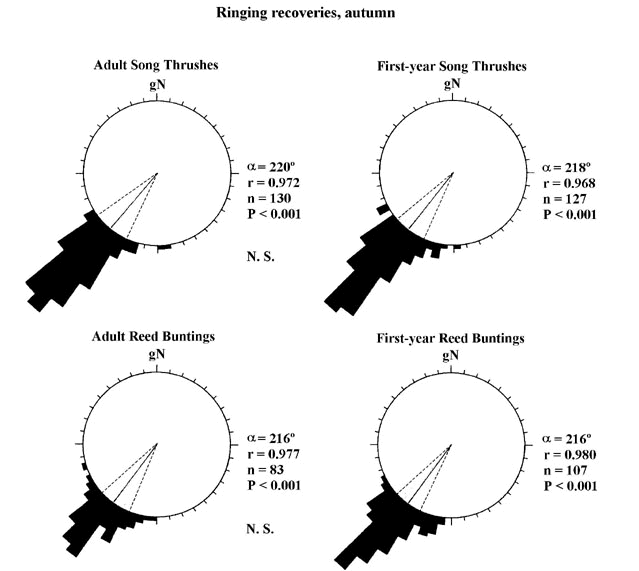
In Fig. 5, we have analysed ringing data for two intracontinental migrants, the Song Thrush Turdus philomelos and the Reed Bunting Emberiza schoeniclus. There were no significant differences in either directional choices or in the concentration of mean orientation between adults and first-year birds. However, if we compare the concentration of headings between adult Pied Flycatchers and Reed Warblers (intercontinental migrants; cf. Fig. 4) with adult Song Thrushes and Reed Buntings, we find that long-distance migrants show significantly more concentrated orientation behaviour (t3>2.8, P<0.001 in all four comparisons according to the 'test for the homogeneity of contration parameters'; Mardia 1972) than do intracontinental migrants (compare Fig. 4 and Fig. 5). Is this an adaptation to compensate for the inevitably higher risks of accumulating orientation errors during long-distance, intercontinental migration flights?
Migratory situation and the integration of different orientation cues
Orientation cage experiments have demonstrated that both diurnal and nocturnal migrants are capable of using directional information from a variety of different environmental sources to select an appropriate compass course for their migratory movements (reviewed by: Moore 1987; Wiltschko and Wiltschko 1991). The orientation behaviour of migrating birds is characterised by complex interrelated mechanisms and flexibility in the use of directional information from the sun, the skylight polarisation patterns, mainly during twilight periods, the rotation of the starry sky and the geomagnetic field (e.g. Baker 1984; Moore 1987; Wiltschko and Wiltschko 1991; Able 1993). The priority and use of different available orientation cues may vary depending on the ecological context within which a migrant finds itself (Sandberg et al. 1988a, b; Sandberg 1991; Pettersson et al. 1991; Åkesson 1993; Sandberg and Moore 1996).
For example, Sandberg et al. (1988a) tested Robins which were captured at two nearby stopover sites in southern Sweden (Falsterbo: 55° 23'N, 12° 50'E and Ottenby: 56° 12'N, 16° 24'E) and showed that birds which arrived on the south-westernmost coast of Sweden (Falsterbo), by overland migration, were lean and temporarily reoriented northwards in unmanipulated orientation cage tests rather than in the expected seasonally appropriate southwesterly direction, which would have brought them across the Baltic Sea. In contrast, Robins that interrupted their flight across the Baltic Sea (presumably originating from Finland) to stopover at Ottenby, on an island in southeastern Sweden, had on average significantly larger fat stores than Falsterbo birds and oriented their migratory activity in the expected southwesterly direction (see also Moore and Kerlinger 1991). In addition to the distinct differences in behaviour under normal geomagnetic field conditions, Ottenby Robins responded to experimental deflections of the geomagnetic field, whereas Falsterbo birds failed to do so, thus indicating a context dependent use of different available orientation cues.
These results have been confirmed in a recent study on the orientation behaviour of Red-eyed Vireos which seemed to integrate geomagnetic information more readily if they were physiologically prepared to cross the Gulf of Mexico (adequate fat stores for a barrier crossing), most likely as a hedge against the possibility of encountering adverse weather like complete overcast and fog while en route (Sandberg and Moore 1996). Considering these results, we argue that migrants are especially sensitive to the availability of directional information when confronted with ecological barriers because of the dire consequences that may occur when landing possibilities are restricted by inhospitable terrain (se also Alerstam 1988, 1990).
DISCUSSION AND CONCLUDING REMARKS
We have attempted to, based on available experimental evidence, bring to the attention the importance of ecological factors in the orientation system of migratory birds. Although endogenous mechanisms are thought to control several aspects of a migrants' physiology and behaviour (e.g. Berthold 1990) such as: the circannual pattern of body mass change, including fat deposition, onset and cessation of migratory activity and seasonally appropriate orientation, it is by no means precluded that external factors may modify migratory behaviour in important ways.
In a number of recent studies, it has been shown that the amount of stored fat play an important role when migrants decide whether or not to embark on migratory flights (e.g. Sandberg et al. 1991; Sandberg and Moore 1996). Furthermore, fat stores can be used to predict whether an individual migrant will select a seasonally appropriate migratory direction or engage in reoriented movements (e.g. Sandberg 1994; Bäckman et al. 1997; Sandberg et al. 1998). Reoriented migration is especially conspicuous during autumn migration when landbirds encounter an ecological barrier for the first time and highlights the complicated web of interactions between energetic status and ecological context in the orientation system of intercontinental migrants.
Not only do birds have to be adequately prepared in terms of energy stores, but long-distance, nonstop flights across ecological barriers pose several challenges to their orientation capacity. We argue that migrants are especially sensitive to the availability of directional information when about to cross an ecological barrier. Several studies have indicated that landbird migrants integrate geomagnetic information more readily into their orientation system while in the process of negotiating a sea passage (Sandberg et al. 1988a, b; Sandberg and Moore 1996; Bäckman et al. 1997). Since orientation errors may have especially dire consequences during barrier crossings where landing possibilities are restricted or completely absent, geomagnetic information may assume greater importance, most likely as a hedge against the possibility of encountering adverse weather en route (e.g. complete overcast, fog), than during overland migration (see also Alerstam 1988, 1990). Clearly there is a need for comparative field studies at inland and coastal sites to disentangle the relative importance of ecophysiological factors and migratory situation in the migratory orientation system of birds (cf. Sandberg 1991; Ehnbom et al. 1993).
Experienced (old) migrants have repeatedly been shown to orient within significantly more narrow limits (less inter-individual scatter in directional choices) than inexperienced birds (Rabøl 1978; Moore 1984; Sandberg et al. 1991). This indicates that old birds are less likely to make orientation errors. The fact that first-year migrants show a higher variability in directional choices suggests either that stabilising selection operates to maintain the orientation within narrow limits (cf. Rabøl 1978), or that adults orient with higher precision due to experience. En route experience, including stopover and winter site attachment, may explain why the orientation performance of adult birds differs fundamentally from that of first-year migrants (see Alerstam 1990).
Given that orientational accuracy differs between experienced and inexperienced migrants, we found it worthwhile to investigate whether there might be a similar difference between long- and short-distance migrants. The rationale behind this being that the longer the migratory distance, the greater the need for orientational precision since even small orientation errors may accumulate in a way that threatens survival. We did indeed find that intracontinental migrants displayed significantly more scatter in their directional choices as compared to intercontinental migrants (but see Åkesson 1993). Admittedly, the number of samples are small, but we find this little used approach interesting and worthy of consideration in future analyses of bird orientation systems.
ACKNOWLEDGEMENTS
This study was supported by grants from the Swedish Natural Science Research Council (NFR) and the Swedish Foundation for International Cooperation in Research and Higher Education (STINT) to R. Sandberg, and from NFR to S. Åkesson.
REFERENCES
Able, K.P. 1977. The orientation of passerine nocturnal migrants following offshore drift. Auk 94: 320-330.
Able, K.P. 1978. Field studies of the orientation cue hierarchy of nocturnal songbird migrants. In: Schmidt-Koenig, K. and Keeton, W. T. (eds) Animal Migration, Navigation and Homing. Heidelberg; Springer-Verlag: 228-238.
Able, K.P. 1980. Evidence on migratory orientation from radar and visual observation: North America. In: Nöhring, R. (ed) Acta XVII International Ornithological Congress. Berlin; Deutsche Ornitologen Gesellschaft: 540-546.
Able, K.P. 1982a. The effects of overcast skies on the orientation of free-flying nocturnal migrants. In: Papi, F. and Wallraff, H. G. (eds) Avian Navigation. Berlin; Springer-Verlag: 38-49.
Able, K.P. 1982b. Field studies of avian nocturnal migratory orientation. I. Interaction of sun, wind and stars as directional cues. Animal Behaviour 30: 761-767.
Able, K.P. 1993. Orientation cues used by migratory birds: a review of cue-conflict experiments. Trends in Ecology & Evolution 8: 367-371.
Able, K.P. & Bingman, V.P. 1987. The development of orientation and navigation behaviour in birds. The Quarterly Review of Biology 62:1-29.
Able, K.P., Bingman, K.P., Kerlinger, P. & Gergits, W. 1982. Field studies of avian nocturnal migratory orientation. II. Experimental manipulation of orientation in White-throated Sparrows (Zonotrichia albicollis) released aloft. Animal Behaviour 30: 768-773.
Åkesson, S. 1993. Effect of geomagnetic field on orientation of the Marsh Warbler, Acrocephalus palustris, in Sweden and Kenya. Animal Behaviour 46: 1157-1167.
Åkesson, S. 1994. Comparative orientation experiments with different species of passerine long-distance migrants: effect of magnetic field manipulation. Animal Behaviour 48: 1379-1393.
Åkesson, S., Karlsson, L. Walinder, G. & Alerstam, T. 1996. Bimodal orientation and the occurrence of temporary reverse bird migration during autumn in South Scandinavia. Behavioural Ecology & Sociobiology 38: 293-302.
Alerstam, T. 1978. Reoriented bird migration in coastal areas: Dispersal to suitable resting grounds? Oikos 30: 405-408.
Alerstam, T. 1981. The course and timing of bird migration. In: Aidley, D. J. (ed) Animal migration. Cambridge; Cambridge University Press: 9-54.
Alerstam, T. 1988. Findings of dead birds drifted ashore reveal catastrophic mortality among early spring migrants, especially Rooks (Corvus frugilegus), over the southern Baltic Sea (in Swedish with English summary). Anser 27: 181-218.
Alerstam, T. 1990. Ecological causes and consequences of bird orientation. Experientia 46: 405-415.
Alerstam, T. & Enckell, P.H. 1979. Unpredictable habitats and evolution of bird migration. Oikos 33: 228-232.
Alerstam, T. & Högstedt, G. 1982. Bird migration and reproduction in relation to habitats for survival and breeding. Ornis Scandinavica 13: 25-37.
Alerstam, T. & Lindström, Å. 1990. Optimal bird migration: the relative importance of time, energy and safety. In: Gwinner, E. (ed) Bird Migration: Physiology and Ecophysiology. Berlin; Springer-Verlag: 331-351.
Bäckman, J., Pettersson, J. & Sandberg, R. 1997. The influence of fat stores on magnetic orientation in day-migrating Chaffinch, Fringilla coelebs. Ethology 103: 247-256.
Baker, R.R. 1984. Bird navigation: the solution of a mystery? London; Hodder & Stoughton: 256pp.
Bairlein, F. 1985. Body weights and fat deposition of Palaearctic passerine migrants in the central Sahara. Oecologia 66: 141-146.
Batschelet, E. 1981. Circular Statistics in Biology. New York; Academic Press: 371pp.
Berthold, P. 1988. The control of migration in European warblers. In: Ouellet, H. (ed) Acta XIX International Ornithological Congress. Ottawa; University of Ottawa Press: 215-272.
Berthold, P. 1990. Genetics of migration. In: Gwinner, E. (ed) Bird Migration: Physiology and Ecophysiology. Berlin; Springer-Verlag: 269-280.
Berthold, P. 1991. Spatiotemporal programmes and genetics of orientation. In: Berthold, P. (ed) Orientation in Birds. Basel; Birkhäuser Verlag: 86-105.
Biebach, H. 1985. Sahara stopover in migratory flycatchers: fat and food affects the time program. Experientia 41: 695-697.
Biebach, H., Friedrich, W. & Heine, G. 1986. Interaction of body mass, fat, foraging and stopover period in trans-Sahara migrating passerine birds. Oecologia 69: 370-379.
Blem, C.R. 1980. The energetics of migration. In: Gauthreaux, S. A., Jr. (ed) Animal Migration, Orientation, and Navigation. New York; Academic Press: 175-224.
Bruderer, B. & Jenni, L. 1988. Strategies of bird migration in the area of the Alps. In: Ouellet, H. (ed) Acta XIX International Ornithological Congress. Ottawa; University of Ottawa Press: 2150-2161.
DeSante, D.F. 1983. Annual variability in the abundance of migrant landbirds on southeast Farallon Island, California. Auk 100: 826-852.
Dolnik, V.R. 1990. Bird migration across arid and mountainous regions of Middle Asia and Kasakhstan. In: Gwinner, E. (ed) Bird Migration: Physiology and Ecophysiology. Berlin; Springer-Verlag: 368-386.
Drury, W.H. & Keith, J.A. 1962. Radar studies of songbird migration in coastal New England. Ibis 104: 449-489.
Ehnbom, S., Karlsson, L., Ylvén, R. & Åkesson, S. A comparison of autumn migration strategies in Robins Erithacus rubecula at a coastal site and an inland site in southern Sweden. Ringing & Migration 14: 84-93.
Emlen, S.T. & Demong, N. 1978. Orientation strategies used by free-flying bird migrants: a radar tracking study. In: Schmidt-Koenig, K. and Keeton, W. T. (eds) Animal Migration, Navigation and Homing. Heidelberg; Springer-Verlag: 283-293.
Emlen, S.T. & Emlen, J.T. 1966. A technique for recording migratory orientation of captive birds. Auk 83: 361-367.
Fransson, T. 1986. The migration and wintering area of Nordic Spotted Flycatcher (In Swedish with English summary). Vår Fågelvärld 45: 5-18.
Gauthreaux, S. A., Jr. 1982. The ecology and evolution of avian migration systems. In: Farner, D. S. & King, J. R. (eds) Avian biology. Vol. 6; London; Academic Press: 93-168.
Hansson, M. & Pettersson, J. 1989. Competition and fat deposition in Goldcrests (Regulus regulus) at a migration stopover site. Vogelwarte 35: 21-31.
Hedenström, A. & Pettersson, J. 1987. Migration routes and wintering areas of Willow Warblers Phylloscopus trochilus (L.) ringed in Fennoscandia. Ornis Fennica 64: 137-143.
Keeton, W.T. 1980. Avian orientation and navigation: new developments in an old mystery. In: Nöhring, R. (ed) Acta XVII International Ornithological Congress. Berlin; Deutsche Ornitologen Gesellschaft: 137-157.
Kettersson, E.D. & Nolan V., Jr. 1985. Intraspecific variation in avian migration patterns. In: Rankin, M. A. (ed) Migration: Mechanisms and Adaptive Significance, Contributions in Marine Science. Vol. 27; Austin; University of Texas Press: 553-579.
Kettersson, E.D. & Nolan V., Jr. 1988. A possible role for experience in the regulation of the timing of bird migration. In: Ouellet, H. (ed) Acta XIX International Ornithological Congress. Ottawa; University of Ottawa Press: 2181-2190.
Kuenzi, A.J., Moore, F.R. & Simons, T.R. Stopover of Neotropical landbird migrants on East Ship Island following trans-Gulf migration. Condor 92: 869-883.
Levey, D. J. & Stiles, F.G. 1992. Evolutionary precursors of long-distance migration: resource availability and movement patterns in Neotropical landbirds. American Naturalist 140: 447-476.
Lindström, Å. 1990. The role of predation risk in stopover habitat selection in migrating Brambling, Fringilla montifringilla. Behavioural Ecology 1: 102-106.
Lindström, Å. & Alerstam, T. 1986. The adaptive significance of reoriented migration of Chaffinches Fringilla coelebs and Bramblings F. montifringilla during autumn in southern Sweden. Behavioural Ecology & Sociobiology 19: 417-424.
Mardia, K.V. 1972. Statistics of directional data. London; Academic Press: 357pp.
Moore, F.R. 1984. Age-dependent variability in the orientation of migratory Savannah Sparrows (Passerculus sandwichensis). Auk 101: 875-880.
Moore, F.R. 1985. Individual variability in the migratory orientation of the Savannah Sparrow, Passerculus sandwichensis. Zeitschrift für Tierpsychologie 67: 144-153.
Moore, F.R. 1987. Sunset and the orientation behaviour of migrating birds. Biological Reviews 62: 65-86.
Moore, F.R. 1994. Resumption of feeding under risk of predation: effect of migratory condition. Animal Behaviour 48: 975-977.
Moore, F.R. & Kerlinger, P. 1987. Stopover and fat deposition by North American Wood-warblers (Parulinae) following spring migration over the Gulf of Mexico. Oecologia 74: 47-54.
Moore, F.R. & Kerlinger, P. 1991. Nocturnality, long-distance migration, and ecological barriers. In: Bell, B. (ed) Acta XX International Ornithological Congress. Wellington; New Zealand Ornithological Congress Trust Board: 1122-1129.
Moore, F.R. & Yong, W. 1991. Intercontinental bird migrants compete for food resources during stopover. Behavioural Ecology & Sociobiology 28: 85-90.
Norberg, U. Vertebrate flight: mechanics, physiology, morphology, ecology and evolution. Berlin; Springer-Verlag: 291pp.
Pennycuick, C. 1989. Bird flight performance: a practical calculation manual. Oxford; Oxford University Press: 153pp.
Pettersson, J. & Hasselquist, D. 1985. Fat deposition and migratory capacity of Robins Erithacus rubecula and Goldcrests Regulus regulus at Ottenby, Sweden. Ringing and Migration 6: 66-76.
Pettersson, J., Sandberg, R. & Alerstam, T. 1991. Orientation of Robins, Erithacus rubecula, in a vertical magnetic field. Animal Behaviour 41: 533-536.
Rabøl, J. 1978. One-direction orientation versus goal area navigation in migratory birds. Oikos 30: 216-223.
Rabøl, J. 1985. The moving goal area and the orientation system of migrant birds. Dansk Ornitologisk Forenings Tidsskrift 79: 29-42.
Ralph, C.J. 1978. Disorientation and possible fate of young passerine coastal migrants. Bird-Banding 49: 237-247.
Ralph, C.J. 1981. Age ratios and their possible use in determining autumn routes of passerine migrants. Wilson Bulletin 93: 164-188.
Richardson, W.J. 1978. Timing and amount of bird migration in relation to weather: a review. Oikos 30: 224-272.
Richardson, W.J. 1990. Timing of bird migration in relation to weather: updated review. In: Gwinner, E. (ed) Bird Migration: Physiology and Ecophysiology. Berlin; Springer-Verlag: 78-101.
Safriel, U.N. & Lavee, D. 1988. Weight changes of cross-desert migrants at an oasis - do energetic considerations alone determine the length of stopover? Oecologia 76: 611-619.
Sandberg, R. 1990. Celestial and magnetic orientation of migrating birds: field experiments with nocturnal passerine migrants at different sites and latitudes. PhD Thesis, Lund University, Lund, Sweden.
Sandberg, R. 1991. Sunset orientation of Robins, Erithacus rubecula, with different fields of sky vision. Behavioural Ecology & Sociobiology 28: 77-83.
Sandberg, R. 1994. Interaction of body condition and magnetic orientation in autumn migrating Robins, Erithacus rubecula. Animal Behaviour 47: 679-686.
Sandberg, R., Bäckman, J. & Ottosson, U. 1998. Orientation of Snow Buntings (Plectrophenax nivalis) close to the magnetic north pole. The Journal of Experimental Biology 201: 1859-1870.
Sandberg, R. & Moore, F. R. 1996. Migratory orientation of Red-eyed Vireos, Vireo olivaceus, in relation to energetic condition and ecological context. Behavioural Ecology & Sociobiology 39: 1-10.
Sandberg, R. & Pettersson, J. 1996. Magnetic orientation of Snow Buntings (Plectrophenax nivalis), a species breeding in the high arctic: passage migration through temperate-zone areas. The Journal of Experimental Biology 199: 1899-1905.
Sandberg, R., Pettersson, J. & Alerstam, T. 1988a. Why do migrating Robins, Erithacus rubecula, captured at two nearby stop-over sites orient differently? Animal Behaviour 36: 865-876.
Sandberg, R., Pettersson, J. & Alerstam, T. 1988b. Shifted magnetic fields lead to deflected and axial orientation of migrating Robins, Erithacus rubecula, at sunset. Animal Behaviour 36: 877-887.
Sandberg, R., Pettersson, J. & Persson, K. 1991. Migratory orientation of free-flying Robins Erithacus rubecula and Pied Flycatchers Ficedula hypoleuca: release experiments. Ornis Scandinavica 22: 1-11.
Terrill, S.B. 1988. The relative importance of ecological factors in bird migration. In: Ouellet, H. (ed) Acta XIX International Ornithological Congress. Ottawa; University of Ottawa Press: 2181-2190.
Terrill, S.B. & Ohmart, R.D. 1984. Facultative extension of fall migration by Yellow-rumped Warblers (Dendroica coronata). Auk 101: 427-438.
Williams, T.C. & Williams, J.M. 1978. Orientation of transatlantic migrants. In: Schmidt-Koenig, K. and Keeton, W. T. (eds) Animal Migration, Navigation and Homing. Heidelberg; Springer-Verlag: 239-251.
Wiltschko, R. 1991. The role of experience in avian navigation and homing. In: Berthold, P. (ed) Orientation in Birds. Basel; Birkhäuser Verlag: 250-269.
Wiltschko, W. & Wiltschko, R. 1991. Magnetic orientation and celestial cues in migratory orientation. In: Berthold, P. (ed) Orientation in Birds. Basel; Birkhäuser Verlag: 16-37.
Wood, B. 1982. The trans-Saharan spring migration of Yellow Wagtails (Motacilla flava). Journal of Zoology, London 197: 267-283.
Yong, W. & Moore, F.R. Relation between migratory activity and energetic condition among thrushes (Turdinae) following passage across the Gulf of Mexico. Condor 95: 934-943.
Fig. 1. Free-flight release experiments with Robins during the autumn migration period in South Scandinavia showing the relationship between the amount of fat stores carried by released birds (visual fat scoring according to Pettersson and Hasselquist (1985)) and the motivation to migrate. The proportion of successful (birds that took off on migratory flights) and unsuccessful (birds that landed) releases are shown for different fat class categories (fat classes with few data have been pooled). Fat Robins were significantly more likely to embark on migration than were birds with low fuel stores (c 2=17.9, df.=2, P<0.001). Modified after Sandberg et al. 1991.
Fig. 2. Autumn orientation of Snow Buntings under clear sunset skies at Resolute in the Canadian Arctic. Results of orientation cage experiments under unmanipulated clear sky control conditions, subdivided into lean (visually estimated fat classes 0-3; top diagram) and fat (fat classes 4-6; bottom diagram) birds. The mean sunset position is indicated outside each circular diagram. Similarly, the direction towards geographic north (gN=360° ) is shown. Each symbol on the periphery of the diagrams represents the mean heading of one individual. The mean vector (a ) of each sample is illustrated by an arrow surrounded by the 95% confidence interval (shaded). Arrow lengths are proportional to the mean vector length (r) and are drawn relative the radius of the circles (radius=1). Significance levels (P) are according to the Rayleigh test (Batschelet 1981). The mean orientation of lean and fat birds were significantly different according to the 'one-way classification test' (F1,26=21.2, P<0.001; Mardia 1972). Based on Sandberg et al. (1998).
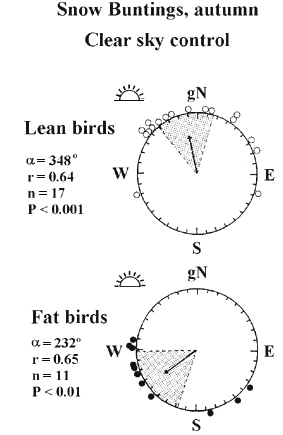
Fig. 3. Vanishing directions of Robins released under clear skies (left diagram) and under complete overcast (right diagram) during autumn migration in South Scandinavia. Wind directions (towards which the wind was blowing) and velocities during the release experiments are shown outside the diagrams, where relevant. The results reveal significantly different orientation behaviour under clear skies and solid overcast, respectively (P<0.001, one-way classification test, Mardia 1972). Other details as in Fig. 2. Modified after Sandberg et al. (1991).
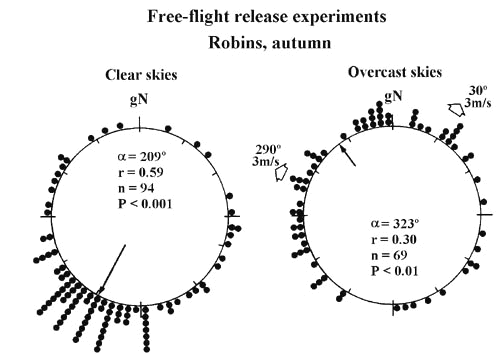
Fig. 4. Circular distributions of ringing recoveries of autumn migrating Pied Flycatchers (top row) and Reed Warblers (bottom row), two intercontinental migrants. Left column shows recoveries of adult birds while the right column shows the corresponding data for first-year migrants. The shortest spoke in each diagram represents the recovery direction of one individual. Pc denotes significance values when the concentration of mean directions were compared between adults and first-year birds (test for the homogeneity of concentration parameters, Mardia 1972). The data is based on annual reports 1972-1986 from the Bird Ringing Centre, Swedish Museum of Natural History. For further details, see Figs 2 and 3.
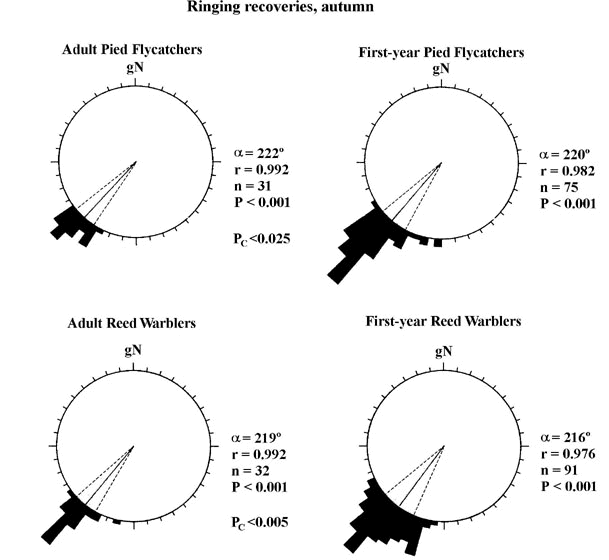
Fig. 5. Circular distributions of ringing recoveries of autumn migrating Song Thrushes (top) and Reed Buntings (bottom), two intracontinental migrants. There were no significant differences in the concentration of mean directions between adults and first-year migrants (N.S.; cf. Fig. 4). For statistical comparisons between adult intercontinental and intracontinental migrants, repectively, see text. Other details as in Fig. 4.