S10.2: Aerodynamics of moult in birds: Effects of wing gaps on flight performance
Anders Hedenström
Department of Animal Ecology Lund University Ecology Building S-223 62 Lund Sweden, fax 46 46 222 47 16, e-mail Anders.Hedenstrom@zooekol.lu.se
Hedenström, A. 1999. Aerodynamics of moult in birds: Effects of wing gaps on flight performance. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban:
536-543. Johannesburg: BirdLife South Africa.The aerodynamics of gaps in the wings of birds that occur periodically during moults has been a neglected field of research. An analysis of the effects of moult gaps in flapping flight is very difficult, but for gliding flight some progress has been made. Flight performance will decrease with increasing gap size (increasing number of missing wing feathers) and gap position (maximum effect in the middle of the wing). Also, the detrimental effects will increase with increasing aspect ratio wings. A few empirical studies have demonstrated that measures of flight performance are reduced due to moult gaps or experimentally induced wing gaps. However, during moult physiological and behavioural changes may also affect flight performance in addition to the purely aerodynamic effects. It is likely that the costs of moult, both in terms of the synthesis of new feathers and reduced flight performance, have moulded the scheduling and sequence of feather replacement in birds. I believe that further research into these questions will be highly rewarding.
INTRODUCTION
During wing moult birds temporarily have gaps in their wings, which are eventually filled by the new feathers at the completion of moult. The fact that the processes of breeding, migration and moult rarely overlap suggests that moult is in some sense costly. But what are these costs? First, there are energetic costs associated with the synthesis of new feather material (reviewed by Lindström et al. 1993), as well as increased costs of thermoregulation due to reduced insulatory capacity of the plumage during moult (e.g. Lustick 1970; Payne 1972). These enhanced energy requirements during moult may increase the foraging rate in exposed situations, which in turn might cause ecological costs such as increased risk of predation. Second, because the missing feathers cause a reduction of wing area and/or wing span it is reasonable to assume that the direct flight costs will increase during moult. However, there is no straightforward way of estimating the aerodynamic costs of moult on the basis of simple momentum jet theory of bird flight (e.g. Pennycuick 1975, 1989), except in cases where the missing feathers cause a reduced wing span. Hence, more sophisticated approaches are required to tackle questions on the effects on flight performance of wing gaps in birds. Such approaches are reasonably tractable for cases of gliding flight performance, but for flapping flight an analysis of the kinematics of wing gaps is still very difficult.
Luckily, there have recently been a few empirical studies on the effects of moult on flight performance to supplement and test the theoretical predictions. In this contribution I will discuss the aerodynamic effects from having gaps in the wings. I will also review the empirical findings of moult and experimentally implemented wing gaps with respect to flight performance in birds. I believe that a deeper understanding of mechanical and functional aspects of moult may help us understand life-history traits such as seasonal timing and speed of moult in birds (cf. Alerstam & Högstedt 1982; Holmgren & Hedenström 1995).
THE MOULT GAP AND WING MORPHOLOGY
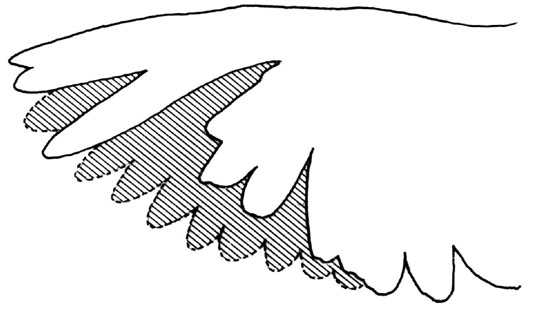
The periodic replacement of the plumage in birds is a process known as moult. During moult wing feathers are missing and therefore the wing area will be reduced. At the stages of moult where the outermost primaries are missing also the wing span will be lower than for a complete wing. Both wing area and wing span are important aerodynamic parameters that affect the aerodynamic performance. In small passerines, the wing area is reduced to about 90% of the pre-moult area during mid-moult, associated with moult score raggedness (missing moult points; Hauikioja 1971) values up to 20 (Hedenström 1998). In Fig. 1 I show the appearance of the wing during typical mid-moult in a passerine. Even larger wing area reductions may occur in hummingbirds, where a female had only 79% of the unimpaired wing area during heavy moult (Chai 1997). During mid-late moult there might be two gaps in the wing of passerines due to simultaneously missing outer primaries and secondaries. Also in large birds with several moult foci more than one gap occur simultaneously on the same wing. In very large birds, such as birds of prey, the relative wing area reduction is usually low and moult is a rather slow process. However, in many birds of prey the primaries are functioning as aerodynamic slots, which reduce the drag of the wing by spreading the wing tip vorticity vertically (Tucker 1995). When such feathers are missing the aerodynamic effect should be larger compared with missing feathers without this function. Some species, such as swans, geese and ducks, shed all their flight feathers simultaneously and thereby loose so much wing area that the capacity to fly is temporarily lost altogether.
THEORETICAL CALCULATIONS OF FLIGHT PERFORMANCE DURING MOULT
There is no trivial approach to predict the effect of moult gaps on aerodynamic performance in birds. One exception is when the moult affects the outermost primaries and hence reduces the wing span. Then, the momentum-jet theory of bird flight (Pennycuick 1975, 1989) can be used because this theory represents a bird by its wing span and mass only. Clearly, power required to fly will increase with a reduced wing span and the lift to drag ratio will decrease. A reduced lift to drag ratio will also affect the performance of gliding flight so that the sink speed will become higher for a given airspeed. Given everything else equal a reduced wing area automatically gives an increased wing loading and so flight performance measures that are affected by wing loading will also be affected by moult. Aerodynamic performance measures that are negatively affected by moult gaps are the acceleration in the roll plane and minimum turning radius (cf. Hedenström 1992).
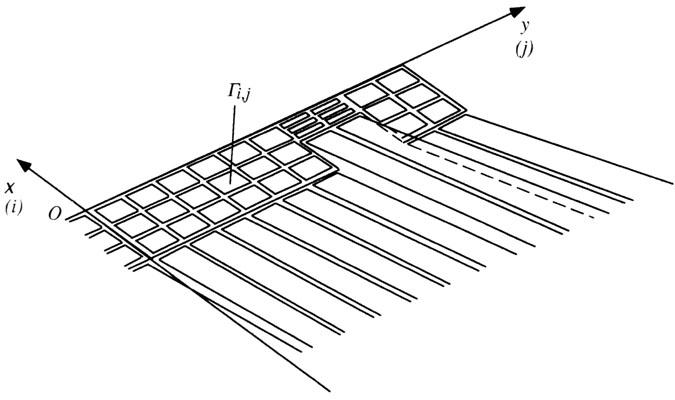
In an attempt to calculate the aerodynamic effects of moult, Hedenström & Sunada (1999) adopted a vortex-lattice (panel) approach from computational fluid dynamics (CFD; Katz & Plotkin 1991; Anderson 1995). The wing and trailing-edge vortices are represented by many small vortex rings as shown in Fig. 2. By integrating (or actually summing) the circulation multiplied by the spanwise extension of the wing vortices and induced velocity the lift coefficient and induced drag coefficient of the trailing vortices can be calculated (Katz & Plotkin 1991). These entities are fundamental when calculating aerodynamic parameters such as lift slope, the effective aspect ratio ARe and the aerodynamic efficiency of the wing (ARe/AR, where AR is the geometric aspect ratio [wing span divided by mean chord]).
Hedenström & Sunanda (1999) used the ideal wing shown in Fig. 2 and modelled the effect of moult gap position, size and the effect of having two simultaneous gaps, which occurs in passerines in the later stages of moult when both outer primaries and secondaries are missing. In general, lift slope and ARe and ARe/AR decreased with the presence of a moult gap (Hedenström & Sunanda 1999). The effect was largest when the moult gap was in the middle of the wing, i.e. inner primaries or outer secondaries are missing. This is because the lift distribution on a wing is near elliptical and so the potential effects are lower towards the wing tip (provided the wing span is unaffected by the gap). Lift slope is reduced with increasing size of the moult gap, while ARe is also reduced but to a lesser extent in large gaps when compared to small gaps. The effect of having two simultaneous gaps was similar to one gap. Finally, the effects of moult gaps on aerodynamic performance increased with increasing aspect ratio. Generally, the effects of moult gaps were rather small, which can be explained by the fact that the detrimental effects from the gaps are to some degree counteracted by the increased aspect ratio of the wing (Hedenström & Sunada 1999).
In conclusion, aerodynamic theory shows that a number of aerodynamic quantities show reduced performance during moult, but overall the expected changes are surprisingly small.
EMPIRICAL STUDIES OF THE EFFECT ON FLIGHT PERFORMANCE DURING MOULT
The direct effects of moult on flight performance have received embarrassingly little attention, but quite recently a few studies have been published. The first, and perhaps most interesting, example refers to Rooks Corvus frugilegus preyed upon by a Peregrine Falco peregrinus (Eutermoser 1961 cited in Ratcliffe 1980). A German falconer, G. Eutermoser, trained a Peregrine to hunt for Rooks and as a control sample he shot a similar number of Rooks that was taken by the falcon. When comparing the two groups, Eutermoser found that among the Rooks taken by the Peregrine 40 % were birds in moult, while only 23 % of the shot sample were so classified. Even if only a tendency, this example shows that moulting birds may be at a higher risk of being predated, presumably because of reduced flight performance.
Tucker (1991) measured the gliding performance of a Harris’ Hawk Parabuteo unicinctus flying in a wind tunnel. The maximum lift to drag ratio dropped from 10.5 before moult to a lowest value of 7.2 during moult when primaries were missing. After moult the lift to drag ratio returned to the pre-moult value of 10.5. Loss of feathers other than primaries did only have minor effects. Tucker (1991) found that of the reduction in lift to drag ratio, reduced wing span and area could only account for 39 % of the total reduction. The remaining reduction was attributed to the fact that the primaries are wing-tip slots that spread the vorticity vertically and thereby reduce induced drag (Tucker 1995). This shows that simple aerodynamic models are not sufficient to estimate the entire suite of aerodynamic effects during moult.
In a completely different approach, Chai (1997) experimentally manipulated the air density by changing the proportion of helium and oxygen and measured the oxygen consumption, stroke amplitude and lift coefficients during natural moult in hovering Ruby-throated Hummingbirds Archilochus colubris. The natural wing gaps were also augmented by trimming the outer primaries and removing the secondaries. In spite of comparatively large reductions of the wing area the lift coefficient changed little, but this was caused by a compensatory reduction of the body mass (Chai 1997). The effect of missing primaries was larger than that of missing secondaries, but this involved a reduced wing span that is of great importance in hovering flight. In further experiments, Chai et al. (1999) found that mass-specific power output declined during moult.
In a series of cage experiments, Swaddle & Witter (1997) measured take-off performance in European Starlings Sturnus vulgaris during natural moult and experimentally induced wing gaps. They found decreased performance of initial escape take-off angle and speed. The Starlings were also flown through a maze to test their manoeuvrability and moulting birds made more contacts with obstacles than the control treatments (Swaddle & Witter 1997).
In conclusion, there is clear empirical evidence that several aspects of flight performance are negatively affected during moult in birds.
ARTIFICIAL WING GAPS AND ECOLOGICAL CONSEQUENCES
Even if there has been little evidence for the aerodynamic consequences of wing gaps, researchers have confidently experimentally reduced wing area and/or span to induce elevated flight costs to study effects on behaviours, survival and breeding performance (reviewed by Mauck & Grubb 1995). For example, these studies have showed effects on the song-flight output in male birds (Møller 1991; Mather & Robertson 1993) and reduced performance of stone-carrying (a sexual display behaviour) in male Black Wheatears Oenanthe leucura (Møller et al. 1995). In a number of studies nestlings show reduced growth rate when their parents were handicapped by removing feathers and sometimes (but not always) the experimental birds themselves show decreased body condition (Mauck & Grubb 1995).
There are problems by using this method for experimentally increasing flight costs in ecological studies. First, the amount of increased cost of flight is not known and so only qualitative effects can be expected. Second, the birds may respond to the experimental manipulation by reducing the amount of flight and hence the total cost of locomotion will not change or change very little. The bird may also respond by adjusting its body mass (Chai 1997; Swaddle & Witter 1997), in which case the total locomotion costs will also remain unchanged. However, there are inescapable costs such as increased risk of predation. In a study of Pied Flycatcher Ficedula hypoleuca, birds with reduced wing areas encountered increased predation levels (Slagsvold & Dale 1996).
DISCUSSION
Obviously there are large gaps in our knowledge about the aerodynamics of moult, although the subject has recently gained some research momentum. Theoretically, we still lack a comprehensive analysis with respect to flapping flight, but for gliding flight the situation is a little better. Independently from the theoretical advancement a few empirical studies have demonstrated that moulting birds experience reduced flight performance compared with non-moulting birds (Tucker 1991; Chai 1997; Swaddle & Witter 1997; Swaddle et al. 1999). However, the theoretical analyses are needed if we are going to fully understand the causes and their relative importance to the total reduction in flight performance observed in experiments or in the field. For instance, Tucker (1991) found that reduction in wing span and area during moult could only partly explain the reduction of lift to drag observed in his Harris’ Hawk, which indicated that additional aerodynamic factors were also involved.
It is not necessarily purely aerodynamic effects that are mirrored in flight experiments. During moult several physiological processes occur in the avian body (Murphy 1996), which may affect metabolism and body mass. At least in some species the main flight muscle Pectoralis major is reduced in size during moult (e.g. Baggott 1975) and this will most certainly affect the power available to aerodynamic work (cf. Pennycuick & Rezende 1984). Hence, the changes in flight performance during moult may be due to aerodynamic, physiological and behavioural factors that may all contribute to the total flight performance of a bird.
An additional factor that could potentially influence flight performance during moult is that the wing gaps are not necessarily perfectly symmetrical between the wings. During feather regrowth the gaps will rather show some degree of fluctuating asymmetry (e.g. Swaddle & Witter 1994, 1998). Such deviations from perfect symmetry should theoretically be associated with reduced flight performance (Thomas 1993) and cage flight experiments have demonstrated that flight performance is indeed detrimentally affected as predicted (Swaddle et al. 1996; Swaddle 1997; Swaddle & Witter 1998).
The prediction that the effect on flight performance will increase with increasing aspect ratio (Hedenström & Sunada 1999) could be of ecological significance. For example, albatrosses have very high aspect ratio wings and their comparatively slow moult is associated with small wing gaps (Prince et al. 1993; Langston & Rohwer 1995), which in turn may be due to elevated flight costs during moult in these birds. Further support of this notion comes from beach-washed dead birds after storms that show an over-representation of moulting birds (Warham 1996).
To conclude, I do believe that an increased aerodynamic understanding of the costs of moult will be very valuable. By way of example, models of the scheduling of annual moults in birds use an overall measure of a loosely defined trait called 'feather quality' (Holmgren & Hedenström 1995). Future models of similar kind should use more accurately defined measures of flight performance that also take into account the present stage of moult for the bird, which should increase the predictive power of such models. It is not unlikely that aerodynamic performance during moult also holds the key to a deeper understanding of timing and moult sequence in birds.
ACKNOWLEDGEMENTS
I am very grateful to Shigeru Sunada for collaborating on flight performance in relation to wing gaps. My research is funded by the Swedish Natural Science research Council.
REFERENCES
Alerstam, T. & Högstedt, G. 1982. Bird migration and reproduction in relation to habitats for survival and breeding. Ornis Scand. 13: 25-37.
Anderson, J.D. Jr. 1995. Computational fluid dynamics. New York; McGraw-Hill: 547pp.
Baggott, G.K. 1975. Moult, flight muscle 'hypertrophy' and premigratory lipid deposition of the juvenile Willow Warbler, Phylloscopus trochilus. J. Zool., Lond. 175: 299-314.
Chai, P. 1997. Hummingbirds hovering energetics during moult of primary flight feathers. J. exp. Biol. 200, 1527-1536.
Chai, P., Altshuler, D.L., Stephens, D.B. & Dillon, M.E. 1999. Maximal horizontal flight performance of hummingbirds: effects of body mass and molt. Physiol. Zool. In press.
Hauikoja, E. 1971. Processing moult card data with reference to the Chaffinch Fringilla coelebs. Ornis Fenn. 48, 25-32.
Hedenström, A. 1992. Flight performance in relation to fuel load in birds. J. theor. Biol. 158: 535-537.
Hedenström, A. 1998. The relationship between wing area and raggedness during molt in the Willow Warbler and other passerines. J. Field. Ornithol. 69, 103-108.
Hedenström, A. & Sunada, S. 1999. On the aerodynamics of moult gaps in birds. J. Exp. Biol. 202: 67 - 76.
Holmgren, N. & Hedenström, A. 1995. The scheduling of molt in migratory birds. Evol. Ecol. 9: 354-368.
Katz, J. & Plotkin, A. 1991. Low-speed aerodynamics: from wing theory to panel methods. McGraw-Hill, New York: 632pp.
Langston, N. E. & Rohwer, S. 1995. Unusual patterns of primary molt in Laysan and Black-footed Albatrosses. Condor 97: 1-19.
Lindström, Å., Visser, G.H. & Daan, S. 1993. The energetic cost of feather synthesis is proportional to basal metabolic rate. Physiol. Zool. 66: 490- 510.
Lustick, S. 1970. Energy requirements of molt in cowbirds. Auk 87: 742-746.
Mather, M.H. & Robertson, R.J. 1993. Honest advertisement in flight displays of Bobolinks (Dolichonyx oryzivorus). Auk 109: 869-873.
Mauck, R.A. & Grubb, T.C. Jr. 1995. Petrel parents shunt all experimentally increased reproductive costs to their offspring. Anim. Behav. 49: 999-1008.
Møller, A. P. 1991. Influence of wing and tail morphology on the duration of song flight in Skylarks. Behav. Ecol. Sociobiol. 28: 309-314.
Møller, A.P., Lindén, M., Soler, J.J., Soler, M. & Moreno, J. 1995. Morphological adaptations to an extreme sexual display, stone-carrying in the Black Wheatear, Oenanthe leucura. Behav. Ecol. 6: 368-375.
Murphy, M.E. 1996. Energetics and nutrition of molt. In: Carey, C. (ed.). Avian energetics and nutritional ecology. New York; Chapman & Hall: 158-198.
Payne, R.B. 1972. Mechanisms and control of molt. In: Farner, D. S., King, J. R. & Parkes, K. C. (eds), Avian Biology Vol. 2; New York: Academic Press: 103-155.
Pennycuick, C.J. 1975. Mechanics of flight. In: Farner, D. S., King, J. R. & Parkes, K. C. (eds), Avian Biology Vol. 5; New York: Academic Press: 1-75.
Pennycuick, C.J. 1989. Bird flight performance: a practical calculation manual. Oxford; Oxford University Press: 153pp.
Pennycuick, C.J. & Rezende, M.A. 1984. The specific power output of aerobic muscle, related to the power density of mitochondria. J. exp. Biol. 108: 377-392.
Prince, P.A., Rodwell, S., Jones, M. & Rothery, P. 1993. Moult in Black-browed and Grey-headed Albatrosses Diomedea melanophris and D. chysostoma. Ibis 135: 121-131.
Ratcliffe, D. 1980. The peregrine falcon. Calton; T & A D Poyser: 416pp.
Slagsvold, T. & Dale, S. 1996. Disappearance of female Pied Flycatchers in relation to breeding stage and experimentally induced molt. Ecology 77: 462-471.
Swaddle, J.P. 1997. Within-individual changes in developmental stability affect flight performance. Behav. Ecol. 8: 601-604.
Swaddle, J.P., Williams, E.V. & Rayner, J.M.V. 1999. The effect of simulated flight feather moult on escape take-off performance in Starlings. J Avian Biol. 30: in press.
Swaddle, J.P. & Witter, M.S. 1994. Food, feathers and fluctuating asymmetry. Proc. R. Soc. Lond. B 255: 147-152.
Swaddle, J.P. & Witter, M.S. 1997. The effects of molt on the flight performance, body mass, and behaviour of European Starlings (Sturnus vulgaris): an experimental approach. Can. J. Zool. 75: 1135-1146.
Swaddle, J.P. & Witter, M.S. 1998. Cluttered habitats reduce wing asymmetry and increase flight performance in European Starlings. Behav. Ecol. Sociobiol. 42: 281-287.
Swaddle, J.P. & Witter, M.S., Cuthill, I.C., Budden, A. & McCowen, P. 1996. Plumage condition affects flight performance in Common Starlings: implications for developmental homeostasis, abrasion and moult. J. Avian. Biol. 27: 103-11.
Thomas, A.L.R. 1993. The aerodynamic costs of asymmetry in the wings and tails of birds: asymmetric birds can’t fly round tight corners. Proc. R. Soc. Lond. B 254: 181-189.
Tucker, V.A. 1991. The effect of molting on the gliding performance of a Harris’ Hawk (Parabuteo unicinctus). Auk 108: 108-113.
Tucker, V.A. 1995. Drag reduction by wingtip slots in a gliding Harris’ Hawk, Parabuteo unicinctus. J. exp. Biol. 198: 775-781.
Warham, J. 1996. The behaviour, population biology and physiology of the petrels. London; Academic Press: 613pp.
Fig. 1. The appearance of the wing during moult in a Yellow Wagtail Motacilla flava, northern Sweden. This individual showed a moult wing area of 88% of the pre-moult wing area associated with a primary score of 15 and a raggedness score of 15 (indicating the missing moult score points – a measure of the wing gap). Stippled area indicates the missing wing area due to moult. Note that when calculating the total wing area a bird for aerodynamic purposes the area of the body between the wings is included.
Fig. 2. Nomenclature for the vortex ring model of the wing and trailing vortices used by Hedenström & Sunanda (1999) to model the aerodynamic effects of moult gaps. Ai,j is the circulation on vortex (i, j), x is the chordwise direction and y is the spanwise direction. It is also shown how a gap in the wing was modelled.