S07.4: Individual variation in renesting intervals, daily energy expenditure and reproductive success in passerines
Kimberly A. Sullivan1, James J. Roper1 & Wesley W. Weathers2
1Department of Biology, Utah State University, Logan UT 84322-5305 USA, fax 435 797 1575, e-mail yejunco@cc.usu.edu; 2Department of Avian Sciences, University of California, Davis CA 95616-8532, USA
Sullivan, K. A., Roper, J. J. & Weathers, W. W. 1999. Individual variation in renesting intervals, daily energy expenditure and reproductive success in passerines. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 401-411. Johannesburg: BirdLife South Africa.
Ornithologists have long recognised that variation in clutch size is an important reproductive tactic in optimising lifetime reproductive success and accordingly, have conducted numerous studies on this topic. Potentially as important, but much less studied, is individual variation in the spacing of reproductive attempts within a breeding season. Using a simulation exercise, we examined the potential effect of variation in renesting interval on annual reproductive success. Breeding season length and nest survival rate both influenced the effect of renesting interval on annual reproduction and under some combinations of breeding season length and nest success rate, a standardised one unit variation in renesting interval had as large an impact on the number of successful nests in a population as a standardised one unit variation in daily nest success rates. We examined the relationship among renesting interval, number of nests attempted, nesting success, fledgling survival and female local survival in a population of Yellow-eyed Juncos Junco phaeonotus and compared these relationships with data available on other species. Female juncos that bred in multiple years initiated more nests year-1, renested faster after a successful nesting attempt, produced more fledglings over the period of this study and had higher survival among their fledged young than those that bred in a single season. We used the doubly- labelled water technique to measure daily energy expenditure in breeding female Yellow-eyed Juncos. Females with relatively low daily energy expenditure after accounting for body mass, air temperature and reproductive stage when measured renested faster and were more likely to breed in multiple years. Data on the demography and daily energy expenditure of Yellow-eyed Juncos combined with data on other avian species suggest that individual females vary in their renesting intervals and propensity to overlap clutches, that this variation is a function of differential access to resources or the ability to use the available resources and that this variation can be an important component of lifetime reproductive success.
INTRODUCTION
In an effort to understand the array of avian life history strategies and the selective forces shaping these strategies, theoretical, observation-based and experimental studies have been carried out addressing the patterns of energy allocation to self-maintenance versus reproduction and among individual offspring (Lack 1968; Ricklefs 1970; Sterns 1992). Birds can vary how they allocate energy to reproduction by varying clutch size, the number of nests attempted and the timing of nest attempts (Burley 1980). One strategy, varying clutch size, has received most of the attention by ornithologists, but over the years, authors have persuasively argued that varying the timing of reproduction potentially can be as important in determining lifetime reproductive success as varying clutch size (Burley 1980; Verhulst et al. 1997).
Here we take three approaches to examining the role of variation in renesting interval on reproductive success. First, we present the results of a simulation exercise which examines how variation in renesting interval and nesting success affect annual reproductive success in a generalised passerine population. Second, we examine data from a population of Yellow-eyed Juncos Junco phaeonotus looking at the relationships among renesting interval, the number of nests attempted, reproductive success and overwinter survival. Third, we examine the possible energetic basis for observed relationships among renesting intervals, reproductive success and survival using data on daily energy expenditure in breeding female birds, as measured using the doubly-labelled water technique.
EFFECT OF VARIATION IN RENESTING INTERVALS ON ANNUAL REPRODUCTIVE SUCCESS IN A SIMULATED PASSERINE POPULATION
As part of a study on the potential demographic effects of increased predation rates on ground nesting birds, two of the authors (J. Roper & K. Sullivan, unpublished data) used Monte Carlo simulations to examine how variation around daily nest survival rate and renesting intervals affected annual reproductive success. In the simulation, populations bred in areas with short (45 day) or long (90 day) breeding seasons and faced a range of either high (the probability of a nest surviving a single day is 0.96, 0.97, 0.98, or 0.99 ) or low (the probability of a nest surviving a single day is 0.91, 0.92, 0.93, 0.94) daily nest survival rates. Small differences in daily nest survival rates translate into large differences in the probability that a nest fledges young. The daily nest survival rates used here represent populations that fledge young from 33-76% of nesting attempts (low predation/high success) and those that fledge young from only 8-19% of nesting attempts (high predation/low success). We assigned our generalised passerine a 27 day nesting period which included nest construction, egg laying, incubation and care of nestlings until fledging based on reported values for small, open-cup nesting passerines with a clutch size of four eggs (Ehrlich et al. 1988).
We generated a uniform random variate (0 < URV < 1) for each day of the nesting period. Each day the nest survived if the URV was less than the assigned daily nest survival rate. This continued until the nest failed or fledged young. A range of renesting intervals (0, 5, or 10 days after failure or 5, 10 or 15 days after fledging young) was added to each nesting attempt and the simulation was then iterated. The short renesting intervals following fledging were selected, because it is not uncommon in passerines for females to initiate a new clutch while their mate continues to care for the previous brood (Burley 1980; Verhulst et al. 1997). When the sum of days expended on breeding equalled the number of days in the breeding season minus the number of days needed for one more successful attempt (18 days for short seasons and 63 days for long seasons) the simulation was terminated. We ran 250 replicates, with each replicate representing a breeding pair, for each combination of independent variables.
For analysis, we log transformed the dependent variable (number of nests fledging young) for each combination of independent variables. In order to compare variables with very different units of measure (such as daily nest survival rate and renesting interval), we standardised the variables using the Standard procedure in SAS. Means were set to 0 and variances to 1. This approach is successful only if the initial variance is biologically meaningful for each independent variable. To illustrate the relative effect of changing each independent variable, we back transformed the data to show the change in the number of nests fledging young for a population of 250 pairs, given a standardised one unit change in the independent variable.
In our simulation, the effects of variation in daily nest survival rate and renesting intervals on annual reproductive success were dependent on breeding season length and the nest survival rate, with the effects being most dramatic when breeding seasons were long and nesting success was high. Under two conditions, a standardised unit change in renesting interval and a standardised unit change in daily nest survival rate had almost identical effects on annual reproduction (Table 1). When the breeding season was short and nest success was high (daily nest survival rates ranged from 0.96 to 0.99), a standardised unit decrease in the renesting interval following fledging (i.e. from 5 to 0 days) had the same effect of increasing the number of nests fledging young in the population by 0.2 nests pair-1 as a standardised unit increase in daily nest survival rate ( i.e. from 0.97 to 0.98). When the breeding season was long and nest success was low, a standardised unit decrease in renesting interval following nest failure had the same effect of increasing the number of nests fledging young in the population by 0.2 nests pair-1 as a standardised unit increase in daily nest survival rate. These results illustrate the potentially large effect variation in renesting interval may have on annual reproductive success.
Although variation in renesting interval had as large an effect on annual reproduction as variation in nest survival rate under some conditions, the potential costs to the breeding pair are very different. In the scenario with short breeding seasons and high nesting success rates, pairs initiated on average 0.3 additional nests when they decreased their renesting interval. In contrast, they initiated 0.1 fewer nests when the daily nest survival rate increased. Under conditions of long breeding seasons and low nesting success rates, pairs initiate on average 1.4 additional nests when they decreased their renesting interval while they initiated 0.2 fewer nests when daily nest survival rates increased. Even if egg production itself may not require exceptional amounts of energy (Perrins 1996; Ward 1996), additional nesting attempts require the expenditure of energy on such activities as nest construction, incubation, and the care of nestlings. Under some conditions, this investment in additional nesting attempts may negatively affect an individual’s condition, the likelihood of surviving until the next breeding season and the survival and future reproductive success of young from previous clutches (Kluyver et al. 1977; Dobson 1990; Verhulst 1998).
RENESTING INTERVALS, NUMBER OF NESTING ATTEMPTS AND REPRODUCTIVE SUCCESS OF YELLOW-EYED JUNCOS
The results of the simulation exercise point out the potentially large role that variation in renesting intervals may have on reproductive success, especially in populations with high nesting success and long breeding seasons. Our database on the breeding activities of a population of Yellow-eyed Juncos provided the opportunity to examine the relationship between variation in renesting intervals and the subsequent number of nests attempted in a breeding season, with reproductive success and survival (K. Sullivan & J. Roper, unpublished data). In this population, the breeding season has been sufficiently long (95, 93, 77, 71 and 52 days for years with reliable first and last egg dates) for some pairs to have fledged 2 or 3 clutches. Nest success rates in most years were in the high portion of the range of the simulation model; varying from 65% to 30% success across years.
A population of Yellow-eyed Juncos in the Chiricahua Mountains of southeastern Arizona were individually marked and monitored from 1984-1991 (see Sullivan 1988, 1989; Sullivan & Roper 1996 for details of the study area, methods employed and breeding biology of this population). Yellow-eyed Juncos are apparently monogamous, ground-nesting, passerines that maintain an all-purpose breeding territory (Moore 1972). They are open cup nesters with a modal clutch size of four eggs. During the 1984-1990 breeding seasons, we individually marked and monitored the nesting attempts of 166 female juncos. In the Chiricahua Mountains, Yellow-eyed Juncos are altitudinal migrants (Horvath & Sullivan 1988) and show site fidelity between breeding seasons. Female between year dispersal distances ranged from 2 to 365 meters (x = 117, SE = 13, n = 63). Each year all suitable habitat within several kms of the main study area was searched multiple times for marked juncos using playbacks of male songs, female chips and nestling distress calls. When a bird failed to return to its previous breeding territory and could not be found on any searches, we assumed that the bird did not survive the winter. It is possible that some females dispersed to outlying areas, but were missed during searches, or dispersed outside the area we searched.
We were concerned that not all territories were visited on a daily or near daily basis and therefore some females may have had fewer of their nesting attempts discovered and also have been less likely to be seen during a breeding season. We divided the marked pairs into four categories based on the frequency of visits to the area where they nested. Territories in the first two categories were visited on a daily or almost daily basis and we found no significant differences in the probability of seeing a bird during the breeding season or the frequency of nesting attempts in those two areas. Territories in the third category were visited at least once a week and females in this area were significantly less likely to breed in multiple years (x2 = 5.96, P < 0.02). Territories in the fourth category were visited at irregular intervals. We dropped the 21 pairs in the third and fourth category from all analyses.
As only a subset of the birds in this study were banded as immatures and some banded birds were still alive at the end of the study, we did not attempt to calculate lifetime reproductive success. Instead we examined annual reproductive success and total reproductive success over the period of this study. Consistent with many studies of lifetime reproductive success (Newton 1989), the total number of young fledged during this study was largely a function of the number of years a female bred at the study site (ANOVA F = 25.43, df = 6,131, P < 0.01; Fig. 1). Only 12% of the females that bred in a single season had one or more offspring recruited into the local breeding population while 44% of the females that bred in multiple seasons had at least one offspring recruited into the breeding population (x2 =10.28, P < 0.01). In contrast, variation in clutch size, 1 to 5 eggs, was not strongly associated with the number of young fledged in a single season or whether an individual bred in multiple years.
Female Yellow-eyed Juncos that bred in multiple years not only had more time available for nesting attempts (number of seasons) but also initiated nests at a higher frequency during each breeding season (ANOVA F = 47.46, df = 6,131, P < 0.01; Fig. 2). Studies on the relationship between number of young raised and overwinter survival have yielded mixed results with some studies reporting a negative relationship (supporting a cost of reproduction) and others reporting a positive relationship (representing variation in female or territory quality, reviewed in Lessells 1991; Moreno et al. 1995; Verhulst 1998). The higher frequency of nesting attempts we detected among multiple year breeders was not a function of increased female experience. Inexperienced pairs of Rock Doves Columba livia had longer renesting intervals than experienced pairs (Burley 1980). In contrast, female juncos that bred in multiple years were more likely to engage in multiple nesting attempts during their first year breeding at our study site than those that bred in only one year (x2 = 18.2, P < 0.01). We found no significant variation in the renesting interval following nest failure (days from nest failure to the first egg of the next nest) between females that bred in multiple years (x = 4.9 days, SE = 0.9, n = 19 females) and those that bred in only one year (x = 7.0 days, SE = 1.6, n = 10 females; t = 1.3 P > 0.20). However, females that bred in multiple years (x = 10.4 days, SE = 1.1, n = 28 females) renested significantly faster after successfully fledging young than those that bred in only one year (x = 15.6 days, SE = 2.8, n = 11; t = 2.1, P < 0.04).
While renesting interval and the subsequent number of nests attempted varied with the number of years a female bred at the study site, nesting success showed little variation among females. We were unable to find significant relationships between nest success as measured by Mayfield estimates of daily nest success, or hatching success or fledging success and the following factors: female experience, whether females bred in one or multiple years, time of the season, or clutch size (all P values > 0.10). In this study, nest failure appeared to be primarily due to predation (Sullivan 1989) and had little apparent relationship with female or territory quality.
As pairs usually provided 21-28 days of post-fledging care before evicting the fledglings from the parental territory (Sullivan 1988), females breeding in multiple years were more likely to initiate a new clutch while their mate was feeding dependent fledglings. Incubating females were rarely observed feeding dependent fledglings (Sullivan 1988). This overlapping of a fledged brood and a new clutch of eggs makes it possible for many pairs to successfully raise two clutches, and a few pairs three clutches, in a breeding season (Weathers & Sullivan 1989). A potential downside of overlapping clutches is that parental investment in the second clutch may lead to a decreased period of parental care, lower survivorship or reduced reproductive success in the previous brood (Verhulst et al. 1997). In Great Tits (Parus major) removal of the second clutch resulted in a longer period of post-fledging care and higher reproductive success among recruits during their first breeding season (Verhuslt et al. 1997). In addition the female Great Tits had higher body mass during the winter, higher overwinter survival in years with poor food availability and produced more fledglings the following year if the second clutch was removed (Verhuslt 1998).
Post-fledging survival, often an important component in lifetime reproductive success (Newton 1989), varied between female juncos breeding in one versus multiple years. In this population 59.5% of 446 fledglings survived the 3-4 week period until independence with most of the mortality (31.4% of fledged young) occurring within 5 days of fledging (Sullivan & Roper 1996). The percentage of broods in which at least one offspring survived the first 2 weeks post-fledging was higher for females that bred in multiple years (60%) than for those that bred in only one year (43%, x2 = 6.26 P < 0.02). This occurred even though females that bred in multiple years were more likely to renest and cease caring for their fledglings sooner than females that bred in only one year.
The positive association of nesting attempts, offspring production, post-fledging survival and return rates suggest that in this population either females or territories varied in quality. Our results are in congruence with those of Rooneem & Robertson (1997) who showed experimentally that female quality, as indicated by the size of the removed first clutch, was an important factor in whether or not Tree Swallows Tachycineta bicolor attempted another nest. They attributed the propensity to renest to variation among females in their foraging skills and efficiency, and the energy they have available to allocate to reproduction. As an alternative, variation in territory quality may influence the timing of reproduction and survival rates. High quality territories may contain more favorable microclimates, higher levels of insect and arthropod prey and refuges for newly fledged young that enhance both female condition and fledging survival. Great Tits decrease their renesting interval and are more likely to overlap clutches when food resources are abundant (Kluyver et al. 1977). Female Blue Tits (Parus caeruleus; Strauss & Glueck 1995) nesting in high quality, deciduous forest, were in better condition, laid larger clutches at earlier dates in the breeding season and had higher nestling survival during the period of peak energy demands than those nesting in poorer quality habitat.
ENERGETIC CORRELATES OF VARIATION IN RENESTING INTERVALS
Examining the variation in energy expenditure among individuals provides one means of identifying the selective pressures shaping patterns of energy allocation (Tinbergen & Dietz 1994). A number of studies have successfully used the doubly-labelled water technique to measure daily energy expenditure (DEE) in breeding birds. Most of these studies have measured DEE in parent birds feeding natural or manipulated broods to address questions concerning the cost of reproduction and evolution of clutch size (for examples see: Bryant 1988; Weathers & Sullivan 1989; Bryant & Tatner 1991; Tinbergen & Dietz 1994; Moreno et al. 1995; Deerenberg et al. 1995; Jonsson et al. 1996). Although many of these studies have uncovered relationships between parental energy expenditure and variables such as: food delivery rate, brood size and parental or nestling condition, these relationships vary greatly among species and between the sexes. Consequently, the nature of the relationship between energy expenditure and fitness remains unknown for most species. In addition, few studies measuring energy expenditure address questions concerning the timing of reproduction and the intervals between attempts. Burley (1980) proposed that by overlapping the care of relatively small clutches instead of producing a single large clutch, parents can reduce the number of young at the stage of peak energy demand and buffer themselves and their offspring from environmental variability. As maximising lifetime reproductive success involves tradeoffs concerning clutch size and the spacing of nesting attempts, the paucity of available information on the energetic basis for decisions on clutch overlap and renesting intervals makes it difficult to interpret the existing data on the evolution of clutch size.
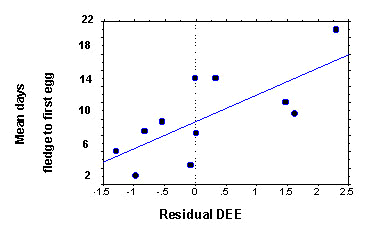
DEE was measured in female Yellow-eyed Juncos in 1985 and 1986 (Weathers & Sullivan 1989) and again in 1988 and 1989 (Weathers, Sullivan & Gumbart, unpublished data) using the doubly-labelled water technique. Because DEE varied as a function of body mass (g0.67), mean air temperature and reproductive stage (incubation, feeding nestlings and feeding fledglings), we used the residual DEE after accounting for these factors in our analyses (Weathers and Sullivan, unpublished data). We used the average residual value for females with multiple measurements.
Among female juncos, the mean renesting interval (days between fledging one brood and laying the first egg of the next clutch) after a successful nesting attempt was positively associated with residual DEE. Residual DEE is the remaining variation resulting after incorporating body mass0.67, mean air temperature and the reproductive stage when DEE was measured into the regression equation. The mean renesting interval = 9.7 + 3.3 residual DEE (r2 = 0.53, n = 11, P < 0.01; Fig. 3). These results support the hypothesis (Rooneem & Robertson 1997) that some females are more efficient at foraging or other activities. Alternatively, variation in DEE and renesting interval could both result from variation in favorable microclimates and prey supplies among territories.
Deerenberg et al. (1995) demonstrated a cost of reproduction in European Kestrels Falco tinnunculus. The DEE of female kestrels, as measured by doubly-labelled water, was positively associated with experimental changes in brood size and negatively associated with survival during the following winter. We failed to find a similar relationship between residual DEE and survival during the following winter for female juncos (x2 = 1.37, P > 0.26). This may be because in contrast to Deerenberg et al.’s study, we did not manipulate many of the females’ broods. However, females that bred in multiple years had lower residual DEE than those that bred in only one season (multiple years x = -0.497, SE = 0.196, n = 9; one year x = .321, SE = 0.32, n = 10; t = -2.064, P < 0.05). The females that bred in multiple years had lower residual DEE even though, as a group, they initiated more nesting attempts, were more likely to overlap parental care of successive clutches and raised more offspring to independence than those that bred in a single season.
It is well recognised among ornithologists that variation in territory quality, experience, and individual differences in foraging efficiency and other parental activities can lead to variation in clutch size among females (Lessells 1991). The costs of reproduction are obscured if high quality females or females on high quality territories exhibit both high levels of reproductive effort and high levels of survival. Consequently, studies examining the costs of reproduction and the evolution of clutch size frequently employ experimental manipulations. In a similar manner, variation among individuals in their access to resources or ability to take advantage of available resources can affect variation in another major reproductive tactic, the timing of nesting attempts (Burley 1980; Rooneem & Robertson 1997; Verhulst 1998). Although variation in nesting intervals has received much less attention than variation in clutch size, under some ecological conditions variation in renesting interval probably has a larger impact on lifetime reproductive success than variation in clutch size. Variation in renesting intervals is likely to be important when breeding seasons are long, there is more than one parent available to care for the young and the decline in the reproductive value of offspring over the season is shallow (Burley 1980; Verhulst et al. 1997). The results of the simulation described here suggests that when nesting success is high the potential for variation in clutch overlap to affect reproductive success is high, while under conditions of low nesting success variation in renesting intervals after nest failures is likely to strongly affect reproductive success. Data from a population of Yellow-eyed Juncos suggest that females vary in their renesting interval with high quality females or females on high quality territories renesting relatively quickly and consequently producing more offspring than other females in the population. Well designed experiments examining patterns of energy allocation, reproductive effort and the costs and benefits associated with the pursuance of different patterns of renesting intervals may measurably increase our understanding of avian life history strategies.
ACKNOWLEDGMENTS
We wish to thank Thomas Gumbart, Joanna Leary and many wonderful research assistants for their invaluable field work, the Southwestern Research Station for logistical support, the U. S. Forest Service, Douglas Ranger District for their cooperation, the USU Berryman Institute for their support and Ken Nagy, Lisa Ballance and an anonymous reviewer for the comments on the manuscript. This work was supported by the National Institutes of Health (HD06552 to K. Sullivan), National Science Foundation (88-02577 to W. Weathers and K. Sullivan), Earthwatch, and the International Federation of Fish and Wildlife Agencies (to M. Conover, T. Messmer and K. Sullivan).
REFERENCES
Bryant, D.M. 1988. Energy expenditure and body mass changes as measures of reproductive costs in birds. Functional Ecology 2: 23-24.
Bryant, D.M. & Tatner, P. 1991. Intraspecies variation in avian energy expenditure: correlates and constraints. Ibis 133: 236-245.
Burley, N. 1980. Clutch overlap and clutch size: alternative and complementary reproductive tactics. American Naturalist 115: 223-246.
Deerenberg, C., Pen, I, Dijkstra, C., Arkles, B., Visser, G.H. & Daan, S. 1995. Parental energy expenditure in relation to manipulated brood size in the European Kestrel Falco tinnunculus. Zoology 99: 39-48.
Dobson, A. 1990. Survival rates and their relationship to life-history traits in some common British Birds. Current Perspectives in Ornithology 7: 115-146.
Ehrlich, R.R., Dobkin, D.S. and Wheye, D. 1988. The Birder’s Handbook. A Field Guide to the Natural History of North American Birds. New York; Fireside Press.
Horvath, E.G. & Sullivan, K.A. 1988. Facultative migration in Yellow-eyed Juncos. Condor 90: 482-484.
Jonsson, K.I., Korpimaki, E. Pen, I. & Tolonen, P. 1996. Daily energy expenditure and short-term reproductive costs in free-ranging Eurasian Kestrels (Falco tinnunulus). Functional Ecology 10: 475-482.
Kluyver, H.N., van Balen, J.H., & Cave, A.J. 1977. The occurrence of time-saving mechanisms in the breeding biology of the Great Tit, Parus major. In: Stonehouse, B. & Perrins, C. (eds.) Evolutionary Ecology. Baltimore; University Park Press: 153-169.
Lack, D. 1968. Ecological Adaptations for Breeding in Birds. London; Methuen.
Lessells, C.M. 1991. The evolution of life histories. In: Krebs, J. R. & Davies, N. B. (eds) Behavioural Ecology An Evolutionary Approach. London; Blackwell Scientific Publications: 32-68.
Moore, N.J. 1972. Ethology of the Mexican Junco (Junco phaeonotus palliatus). Ph.D. dissertation, University of Arizona, Tucson.
Moreno, J., Cowie, R.J., Sanz, J.J. & Williams, R.R. 1995. Differential response by males and females to brood manipulations in the Pied Flycatcher: energy expenditure and nestling diet. Journal of Animal Ecology 64: 721-732.
Newton, I. 1989. Lifetime Reproduction in Birds. Newton, I. (ed) Academic Press, New York.
Perrins, C.M. 1996. Eggs, egg formation and the timing of breeding. Ibis 138: 2-15.
Ricklefs, R.E. 1970. Clutch size in birds: outcome of opposing predator and prey adaptations. Science 168: 599-600.
Rooneem, T.M. & Robertson, R.J. 1997. The potential to lay replacement clutches in Tree Swallows. Condor 99: 228-231.
SAS Institute. 1990. SAS Procedures Guide. Version 6, 3rd Edition. Carey; SAS Institute Inc.: 551-557.
Stearns, S.C. 1992. The Evolution of Life Histories. Oxford; Oxford University Press.
Strauss, M.J. & Glueck, E. 1995. Effects of habitat quality on breeding phenology and reproductive success in Blue Tits (Parus caeruleus). Vogelwarte 38: 10-23.
Sullivan, K.A. 1988. Ontogeny of time-budgets in Yellow-eyed Juncos: adaptations to ecological constraints. Ecology 69: 118-124.
Sullivan, K.A. 1989. Starvation and predation: age specific mortality in juvenile juncos. Journal of Animal Ecology 58: 275-286.
Sullivan, K.A. & Roper, J.J. 1996. Impacts of predation on passerine post-fledging success. Transactions of the 61st North American Wildlife and Natural Resource Conference 61: 77-85.
Tinbergen, J.M. & Dietz, W.M. 1994. Parental energy expenditure during brood rearing in the Great Tit (Parus major) in relation to body mass, temperature, food availability and clutch size. Functional Ecology 8: 563-572.
Verhulst, S., Tinbergen, J.M. & Daan, S. 1997. Multiple breeding in the Great Tit. A trade-off between successive reproductive attempts? Functional Ecology 11: 714-722.
Verhulst, S. 1998. Multiple breeding in the Great Tit, II. The costs of rearing a second clutch. Functional Ecology 12: 132-140.
Ward, S. 1996. Energy expenditure of female Barn Swallows (Hirundo rustica) during egg formation. Physiological Zoology 69: 930-951.
Weathers, W.W. & Sullivan, K.A. 1989. Juvenile foraging proficiency, parental effort, and avian reproductive success. Ecological Monographs 59: 223-246.
Table 1. Effect of variation in daily nest survival rate and renesting interval on annual reproduction. All values are means + 1 SE.
Fig. 1. The total number of young fledged by females observed breeding for periods of 1, 2 and 3 years at our study site (means + 1 SE). Females breeding in only 1 year fledged significantly fewer young than those breeding in 2 years (t = 6.88, P < 0.01) which fledged significantly fewer young than those breeding in 3 years (t = 2.92, P < 0.01 ).
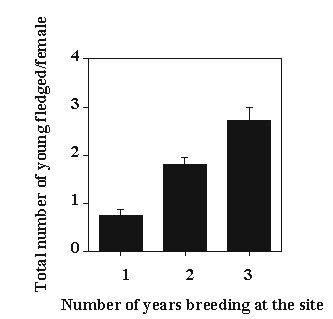
Fig. 2. Frequency of nesting attempts for females observed breeding 1, 2 and 3 years at our study site (mean + 1 SE). Females that bred in 1 year initiated fewer nests per season than those that bred in 2 years ( t = 3.42, P < 0.01). There was no significant difference between attempts year-1 for females breeding in 2 or 3 years (t = 1.62, P > 0.10).
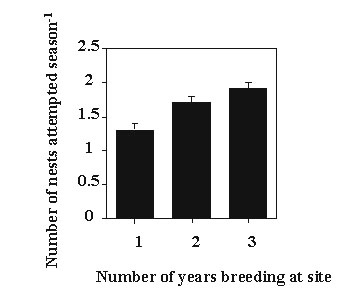
Fig. 3. Relationship between residual daily energy expenditure (DEE), after accounting for mass (g0.67), mean air temperature and reproductive stage, and mean interval between successfully fledging one brood (fledge) and initiating a new clutch (first egg) among female juncos. Mean renesting interval = 9.7 + 3.3 (residual daily energy expenditure), r2 = 0.53, n = 11, P < 0.01. Females with relatively high DEE take longer to renest.