S06.5: Survival rates of understorey forest birds in Peru
Charles M. Francis1, John S. Terborgh2, & John W. Fitzpatrick3
1Bird Studies Canada, PO Box 160, Port Rowan, Ontario, N0E 1M0, Canada, fax 1 519 586 3532, e-mail cfrancis@bsc-eoc.org; 2Center for Tropical Conservation, Duke University, Durham, North Carolina 27708, USA, e-mail manu@acpub.duke.edu; 3Cornell Laboratory of Ornithology, Sapsucker Woods Road, Ithaca, New York, USA, e-mail jwf7@cornell.edu
Francis, C. M., Terborgh, J. S. & Fitzpatrick, J.W. 1999. Survival rates of understorey forest birds in Peru. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 326-335. Johannesburg: BirdLife South Africa.Estimated mean survival rates of Neotropical forest birds vary considerably among recent studies. Some studies found survival rates of tropical birds to be similar to those of temperate species, while others, mainly conducted on islands, found relatively high survival rates of tropical birds. We used mark-recapture analysis with 10 years of mist-netting data from Manu National Park, Peru to estimate survival rates for 14 species of frequently captured understorey birds in the Amazonian rainforest. For most species, models were required that allowed for lower apparent survival rates in the first year after banding compared to later years. Mean apparent survival rate across species from these models was 0.47 in the first year after banding and 0.68 (S.E. 0.02) in subsequent years. This suggests that initial captures include a mix of resident birds and non-resident, transient, or immature birds with lower survival rates and/or site fidelity. Estimated survival rates for residents are comparable to recently published estimates for understorey species from Puerto Rico (mean 0.68) and from Trinidad (mean 0.66), but significantly higher than those from Panama (mean 0.56-0.58). These results indicate that higher survival rates in the tropics are not confined to islands, and suggest that survival rates of understorey birds at one study site in Panama may be different from those elsewhere in the Neotropics.
INTRODUCTION
Tropical birds have long been assumed to have much higher survival rates than temperate birds, but this notion was recently challenged by Karr et al. (1990). These authors pointed out that much of the available evidence was based upon small sample sizes and did not use modern statistical methods. They used modified Cormack-Jolly-Seber mark-recapture models (Pollock et al. 1990, Lebreton et al. 1992) to estimate survival rates for 25 species in Panamanian rainforest, and found that apparent survival rates for these species averaged 0.55, which was very similar to average survival rates for a suite of temperate-breeding passerines estimated with similar methods (0.54).
Subsequently, Faaborg (1995) used similar models to analyse data for tropical residents at a study site on the island of Puerto Rico, and found an average survival rate for nine species of 0.68 - substantially higher than estimates from the Karr et al. (1990) site in Panama. Johnston et al. (1997) analysed survival rates for 17 species on the island of Trinidad, and estimated mean survival rates of 0.65, again significantly higher than estimates from Karr et al. (1990). They found that two ‘age’ class models were required for most species (where ‘age’ refers to years since banding, rather than true age). These allow for lower survival and/or higher emigration for newly banded birds relative to birds returning from previous years. Such models may be required due to the presence of either young birds or transients (non-residents) in the initial captures, which have lower survival rates or lower site fidelity than resident adults. Johnston et al. (1997) suggested that the estimates of Karr et al. (1990) may be biased downwards, because they failed to take into account the potential impact of these transients on their estimates. However, new analyses of the data from Panama (Brawn et al., this volume) continue to indicate that survival rates of birds from this site in Panama are significantly lower than those from either Trinidad or Puerto Rico, even after allowing for transients or young birds. Although methods differed slightly among the studies (for example, the Panama study generally involved two sampling sessions per year, rather than one), the different results suggest that mean survival rates may genuinely differ among tropical American study sites. It may be significant that some of the original studies indicating high survival rates in the Neotropics, particularly those reported by Snow and Lill (1974) from Trinidad and Willis (1974) from Barro Colorado Island in Panama, were also from islands, while the study of Karr et al. (1990) was on mainland Panama.
We analysed capture data for a selection of understorey birds in the Amazonian rainforest of Peru, to determine whether their apparent survival rates more closely approached those observed in Panama, or on the islands of the Caribbean. Data were collected from a 10-year capture-recapture program using mist nets, and were analysed using similar models to those of Johnston et al. (1997), so the results should be directly comparable. Because net lines covered an area of >100 ha, we also investigated movements of birds between capture locations, to explore the possibility that movement of birds away from the initial capture site may bias survival estimates downwards.
METHODS
The study area was in mature floodplain forest, early primary successional forest, and some intervening swamp forest around Cocha Cashu Biological Station in Peru's Manu National Park (11oE54' S, 71oE18' W, elevation ca. 400 m). The study area and its bird community is described in detail elsewhere (Robinson & Terborgh 1990, Terborgh 1990, Terborgh et al. 1990). The area used for the capture-recapture study included most of the census area used by Terborgh et al. (1990) and some additional areas to the east.
Birds were sampled using mist-nets (12 x 2 m) that were set end-to-end in lines of 10 to 40 nets along trails. Nets were opened at dawn, checked regularly through the day, then furled before dusk. In most cases, each line of nets was operated for 3-4 days per year. Birds were processed (banded and measured) and released at the net where they were captured. All captured birds were banded with uniquely numbered bands, and many also received coloured plastic leg bands. In addition, wing length, body mass, age and sex (if they could be distinguished), and the precise net of capture were recorded for each bird.
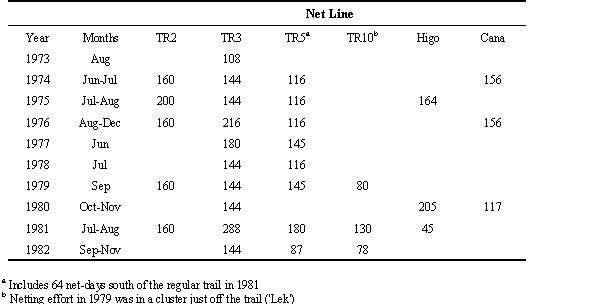
The most consistent netting took place along TR3 (Trail 3), where 36 nets were set starting 100 m north of the base camp on Cocha Cashu, and running approximately northeast for about 430 m (Table 1). In most years, a parallel line of 28 nets was set on TR5 about 300 m east of TR3 (near the edge of the census area of Terborgh et al. 1990), and another parallel line of 40 nets was set on TR2, about 350 m farther east/south-east. Additional net lines run in more than one year included 26 nets on Trail 10, about 300 m north of Trail 3; 41 nets at ‘Higo’ about 600 m south of Trail 3; and 39 nets at ‘Caña’ about 100 m east of Higo. Nets were also set for single netting sessions, usually of 3-4 days each, at several additional locations, of which the farthest was Riachuelo, about 1200 m east, and 800 m north of the base camp, where 19 nets were set for 3 days in 1974.
For the main analysis, we included data from all captures in the study area because many birds banded in one net lane were subsequently recaptured in other net lanes. We selected the 14 species with at least 50 individuals captured during the 10-year interval, of which from 11 to 30% were recaptured at least once in subsequent years (Table 2). We also repeated the analysis using only captures from TR3 and TR5, to determine whether inclusion of data from other transects might bias survival estimates.
For each individual, we created a capture history, consisting of 1's in the years it was captured, and 0's in the years it was not captured. We then used open population capture-recapture analysis methods to estimate survival rates for each species while allowing for the fact that many individuals, even if present in a particular year, may not have been captured that year (Pollock et al. 1990, Lebreton et al. 1992). Because of differences in capture effort among years (Table 1), we assumed capture rates, p, varied among years (t), which we denoted p(t). We also tested models assuming capture rates remained constant every year, but these were rejected in all cases. We then estimated survival rates using two different models. The first assumed that apparent survival rates (f ) of all birds were the same, and remained constant over time, denoted f (c). The second model allowed apparent survival rates for newly banded birds, in their first year after banding, to differ from apparent survival rates in subsequent years, denoted (f )(a2). This 2 ‘age’-class model is mathematically equivalent to the transient models of Pradel et al. (1997), and differs only in the interpretation of apparent survival rates in the first year after banding. In the model of Pradel et al. (1997) the difference between this survival rate, and apparent survival in subsequent years is interpreted as being due to a proportion (g) of the newly banded birds being transients that have zero probability of remaining in the study area and being recaptured in the future. However, inclusion of immature birds, with lower survival or higher emigration rates, or a mixture of birds with high and low capture probabilities will also lead to differences in apparent survival rates between newly banded and previously banded birds. These effects cannot be differentiated without additional information, so we did not attempt to estimate proportions of transients.
Recent work suggests that model selection should be based on Akaike’s Information Criterion, adjusted if necessary for over-dispersion in the data using a quasi-likelihood approach (QAICc; Burnham et al. 1995). This is based upon the principle of trying to select the model with the smallest number of parameters that adequately fits the data. In our results, we have indicated the preferred model based on this criterion, but we nevertheless present estimates derived from both models. We fitted all models using program MARK (White 1998), selecting the option to calculate standard errors for the estimates based on numerical approximations to the second partial derivatives.
We also estimated movement distances for each species between captures. For this analysis, we considered only the location of the first capture for each individual bird in a year. We estimated separately mean distance between first and second year of capture, and mean distances between subsequent years. We calculated approximate co-ordinates for each capture location from a scale map showing the locations of all nets. We then calculated the straight-line distance between the locations to estimate movement distance.
RESULTS
Models allowing apparent survival rates in the first year of capture to differ from those in subsequent years were selected for eight out of the 14 species (Table 1). For all 14 species, mean apparent survival rates over the first year after capture averaged 0.47 (SE 0.04), while those in subsequent years averaged 0.68 (SE 0.02), a difference of 0.21. If the difference between first and subsequent years had been ignored, the mean apparent survival rates would have been estimated as 0.61.
Some of the difference in apparent survival between first and subsequent years could have resulted from inclusion of data from several net lanes that were run in only one year (such that fewer of those birds were likely to be recaptured). To test this, we repeated the analyses using only data for birds captured in TR3 or TR5. The sample size for many species was substantially reduced, leading to reduced precision for many estimates. Nevertheless, mean survival rates across species were virtually unaffected. For the constant survival model, estimated apparent survival rates were 0.63 (S. E. 0.05), while for the 2-age class model, mean survival in the first year was 0.46 (S. E. 0.04) and in subsequent years was 0.71 (S. E. 0.05). The 2-age class model was again preferred for many species.
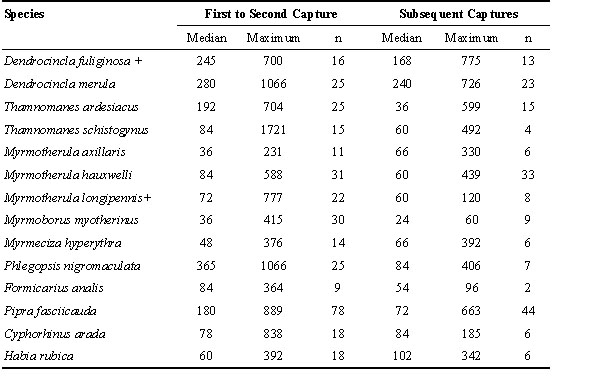
Median movement distances between the first capture location in the first year, and the first location in the next year of capture ranged from 36 m to 365 m (Table 3). As expected, the two species known to be obligate ant followers, Dendrocincla merula and Phlegopsis nigromaculata, had the highest median movement distances. Maximum distances were over 200 m for all species, and 3 species (including both ant followers) had maximum movements over 1 km. The maximum distance between the extremes of the net lines operated most regularly (TR3, TR5, and TR2) was only about 1 km, so longer movements necessarily involved some of the less frequently used net lanes. Median movement distances after the first year ranged from 24 m to 240 m. For 10 of the 14 species, these appeared to be somewhat smaller than movements between the first pair of years, but these estimates may be biased because some of the more distant net lanes were only operated in one year, reducing the potential for detecting long-distance movements in subsequent years.
Estimated apparent survival rates for each species were not significantly correlated with mean body mass (r = 0.4, P = 0.15), mean wing length (r = 0.14, P = 0.6), or mean movement distance in the first year (r = 0.04, P = 0.91) or in subsequent years (r = 0.2, P = 0.49). However, mean movement distances were positively correlated with wing length in the first (r = 0.58, P = 0.03) and in subsequent years (r = 0.70, P = 0.006), and more weakly with body mass (in both cases, r = 0.40, P = 0.15).
DISCUSSION
Mean survival rates of the birds we studied at Manu were similar to those reported by Faaborg and Arendt (1995) for Puerto Rico, and Johnston et al. (1997) for Trinidad. Our data reaffirm the suggestions of those authors that tropical birds do have higher survival rates than temperate species, and show that this difference is not restricted to islands. Our survival estimates are significantly higher than those of Karr et al. (1990) for Panama, suggesting that there may be significant variation in avian demography in different areas of the tropics. The mean body mass of our 14 species was 27.4 g, similar to that of the samples from Trinidad (28.7 g; Johnston et al. 1997) and from Panama (25.1 g; Karr et al. 1990). However, although body size has been found to be generally correlated with survival rates in some comparative studies (Saether 1989), we found no relationship between the two in our study. Johnston et al. (1997) used body mass as a covariate in their comparative analyses, because of significant relationships between body mass and survival estimates for their temperate species, but our analysis of their data also indicate no relationship between survival rate and body mass of their sample of species from Trinidad.
Despite the fact that survival rates from our study are clearly higher than those of many temperate species, as well as those from the Karr et al. (1990) study, our mean survival estimate of 0.68 falls short of the estimates of 0.8-0.9 that were formerly quoted for tropical birds, such as by Snow and Lill (1974) for selected birds in Trinidad, or Fogden (1972) for birds in Sarawak, Malaysia. What could explain these discrepancies?
In general, the higher mean survival rates (>0.8) tend to come from studies involving intensive observations of colour-marked birds, while the lower estimates (<0.7) come from mist-netting studies. These methods differ both in the segment of the population being monitored, and in the method of follow-up, both of which could lead to discrepancies.
The study by Fogden (1972) was based on observations of colour banded birds over a single year of study. Formal capture-recapture models were not necessary because the resighting rate was apparently 1.0 or very close to it. In any case, if any individuals were missed, this should have led to an underestimate of survival rates. His analysis was restricted to pairs of birds observed to be resident in a 20 ha plot. Although it is possible that the year of his study was exceptionally favourable for survival, there is no obvious reason to suspect bias upwards. The study by Snow and Lill (1974) was extended over a 9-year period, and hence was unlikely to have represented an exceptionally favourable time period. They also did not use formal capture-recapture analysis methods, but again their estimates would tend to be biased low if they failed to detect any individuals. Most of their data were for two species of manakins, Pipra erythrocephala and Manacus manacus, and it is possible that these species, especially the adult males displaying on leks (which comprised most of the sample) may have higher survival rates than other tropical passerines. Willis (1974) found survival rates of one species of antbird to be over 80%, though survival for two other species averaged 71%. Greenberg and Gradwohl (1986) initially estimated survival of territorial Checker-throated Antwrens Myrmotherula fulviventris at 0.61, but with additional data revised their estimate upwards to 0.75, suggesting their early estimate may have been biased by band loss (Greenberg and Gradwohl 1997).
In contrast, capture-recapture estimates based on mist-nets are sampling from different segments of the population than these intensive observational studies. Mist nets set along transects, or in other systematic ways through the forest, are likely to capture all components of the population, and may even tend to over-represent floaters, transients, and immature birds if residents have a tendency to become net shy (see below). The models used in the current analysis exclude transients from the adult survival estimates as long as they disappear from the study area after their first capture. However, survival rates of non-territorial floaters that remain in the study area will be averaged with those of residents. If, as suggested by Piper and Oatley (this volume) these birds have lower survival rates than residents, estimated survival rates from capture-recapture models will be intermediate between those of residents and floaters. Similarly, if non-breeding or immature birds remain with the breeding pair for a protracted period (e.g. > 1 year), their apparent survival rates, which may be lower due to greater mortality and/or higher dispersal, will also be averaged with those of residents.
Survival estimates from mist net samples could also potentially be affected by net shyness – a tendency for birds to avoid recapture after their initial capture. One of us (J. Terborgh, unpublished data) tested for net shyness in 1975 by visually estimating the proportion of individuals in the population along TR3 that were already marked from the previous year, prior to carrying out mist-netting along that transect. Across species, the proportion of previously marked individuals captured was approximately one-third less than the proportion in the sample estimated visually. This suggests that at least some marked birds learn to avoid nets, and further more, that this learning persists for at least one year. If the learning is permanent (i.e. does not change over time after the first capture), and if it is the same for all individuals, this will not bias survival estimates (although it would strongly bias capture-recapture estimates of population size). This is because the survival models are conditional on marked birds. However, if some individuals such as territorial resident adults are more likely to learn to avoid nets than others, the survival estimate could be based on a sample biased away from those birds. Further bias could occur if birds become progressively better able to avoid nets after subsequent captures. Although this type of learning has not been demonstrated, it is worth noting that two of the species with particularly low survival estimates are species that move slowly through the understorey by walking (Formicarius analis) or hopping (Myrmoborus myotherinus) on the ground. This behaviour may lend itself to spotting and avoiding mist-nets, especially after one or more captures along a well-worn net line. This could be associated with the fact that Myrmoborus myotherinus was the only species for which the point estimate for apparent survival of previously marked birds was actually lower than that for newly marked birds (Table 2). Although this difference was not statistically significant and hence could be due to chance, it could also be caused by increasing net shyness after subsequent captures.
Finally, it is important to remember that any capture-recapture or mark-resighting study, whether based on observations, or mist-netting, will underestimate survival to the extent that birds emigrate from the study area (and survive). Karr et al. (1990) assumed that dispersal and subsequent survival for tropical birds was virtually nil, but few data are available to back this up. Many tropical bird populations are believed to contain a significant proportion of floaters that presumably range over relatively large areas in search of vacant territories. It would seem quite likely that such individuals, if originally marked within a study area, could settle in a territory outside that study area. Also, for territorial species with permanent pair bonds, few data are available on behaviour of birds that lose a mate. Willis (1974) found interspecific variation in the behaviour of 3 species of antbirds. Female Hylophylax naevioides Spotted Antbird and Phaenostictus mcleannani Ocellated Antbird were more likely than males to change territories if they lost a mate, while both sexes were equally likely to move in Gymnopithys bicolor Bicolored Antbird. He speculated that in his study, about 10-15% of disappearing birds probably had emigrated rather than died. In a mist-netting study, a bird need only move one or two territories away from a net lane to have effectively emigrated.
Our data on movement patterns indicate that some adults move significant distances from their original capture locations. For 10/14 species, some birds known to have been in the study area for >1 year were captured >250 m from the site of their previous capture, suggesting that adults may sometimes switch territories (250 m is approximately the diameter of a 5 ha territory, which is the average for many of these relatively common species; Terborgh et al. 1990). Furthermore, these movement estimates are necessarily low, because nets in the more distant lines were not set every year and because some birds probably moved beyond any of our net lanes. Although we cannot be sure that these birds moved permanently, the data do suggest that it is not appropriate to assume that birds are permanent residents without further examining the issue.
Bias due to emigration will affect not only survival estimates based on mist-net captures, but also those based on mark-resighting, suggesting that any of the published estimates will tend to underestimate survival. However, the bias may be greater for mist-net samples because of the inclusion of additional segments of the population that are more likely to emigrate, such as floaters or non-breeding members of a breeding group.
Interestingly, a study of the genus Turdus in the Western Hemisphere using cross-sectional data on age ratios in museum samples of the population, rather than longitudinal data from capture-recapture analyses, suggests survival rates in the tropics in excess of 0.80 for several species (Ricklefs 1997). Estimates for temperate species in the genus were similar to published estimates for other temperate passerines (Karr et al. 1990). Although such cross-sectional studies require some rather restrictive assumptions (e.g., equal collecting probability of adults and first-year birds, stable age distributions, all birds accurately aged), and are less flexible than longitudinal studies for looking at other aspects of demography, they nevertheless provide independent confirmation of differences in survival rates between temperate and tropical species. Furthermore, this method is not affected by emigration rates, and provides further evidence that survival rates greater than 0.80 may well be more typical of tropical birds than some of the lower estimates derived from capture-recapture estimates.
The general tendency for survival estimates based upon mist-netting data to be lower than those estimated using alternative methods, such as intensive observations of colour-marked birds, suggests that the use of mist-net data may tend to underestimate survival rates, at least for breeding adults. However, in very few cases are data available for the same species using multiple methods, and in no case have data been collected using multiple methods at the same site. As a result, with data presently available, it is not possible to determine whether discrepancies among studies are due to differences in the biology of the species involved or differences in the study methods. This issue stands as an invitation to further investigation, preferably involving longer-term observations of marked individuals at a particular site using both mist-netting and observational sampling methods.
ACKNOWLEDGMENTS
We would like to thank the responsible officials in both the Division of Forestry and Wildlife of the Peruvian Ministry of Agriculture and the administration of the Manu National Park for permission to carry out the study. We also thank the many different people who assisted with the banding, and Cecilia Yun-Lin Fung for assistance in computerising the banding data. Preparation of the manuscript was supported, in part, by Bird Studies Canada. Fieldwork was supported by research grants from the United States National Science Foundation.
REFERENCES
Burnham, K.P., White, G.C. & Anderson, D.R. 1995. Model selection in the analysis of capture-recapture data. Biometrics 51: 888-898.
Faaborg, J. & Arendt, W. 1995. Survival rates of Puerto Rican birds: are islands really different? Auk 112: 503-507.
Fogden, M.P.L. 1972. The seasonality and population dynamics of equatorial forest birds in Sarawak. Ibis 114: 307-343.
Greenberg, R. & Gradwohl, J. 1986. Constant density and stable territoriality in some tropical insectivorous birds. Oecologia 69: 618-625.
Greenberg, R. & Gradwohl, J. 1997. Territoriality, adult survival, and dispersal in the Checker-throated Antwren in Panama. Journal of Avian Biology 28: 103-110.
Johnston, J.P., Peach, W.J., Gregory, R.D. & White, S.A. 1997. Survival rates of tropical and temperate passerines: a Trinidadian perspective. American Naturalist 150: 771-789.
Karr, J.R. 1990. Avian survival rates and the extinction process on Barro Colorado Island, Panama. Conservation Biology 4: 391-397.
Karr, J.R., Nichols, J.D., Klimkiewicz, M.K. & Brawn, J.D. 1990. Survival rates of birds of tropical and temperate forest: will the dogma survive? American Naturalist. 136: 277-291.
Lebreton, J.-D., Burnham, K.P., Clobert, J. & Anderson, D.R. 1992. Modelling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecological Monographs 62: 67-118.
Martin, T.E. 1996. Life history evolution in tropical and south temperate birds: what do we really know? Journal of Avian Biology 27: 1-10.
Pollock, K.H., Nichols, J.D., Brownie, C. & Hines, J.E. 1990. Statistical inference for capture-recapture experiments. Wildlife Monographs 107: 1-97.
Pradel, R., Hines, J.E., Lebreton, J.-D. & Nichols, J.D. 1997. Capture-recapture survival models taking account of transients. Biometrics 53: 60-72.
Ricklefs, R.E. 1997. Comparative demography of New World populations of thrushes (Turdus spp.). Ecological Monographs 67: 23-43.
Robinson, S.K. & Terborgh, J.S. 1990. Bird communities of the Cocha Cashu biological station in Amazonian Peru. In: Gentry, A.H. (ed.) Four Neotropical Rainforests, New Haven, USA: Yale University Press: 199-216.
Saether, B.E. 1989. Survival rates in relation to body weight in European birds. Ornis Scandinavica 20: 13-21.
Terborgh, J.S. 1990. An overview of research at Cocha Cashu Biological Station. In: Gentry, A.H. (ed.) Four Neotropical Rainforests, New Haven, USA: Yale University Press: 48-59.
Terborgh, J.S., Robinson, S.K., Parker III, T.A., Munn, C.A. & Pierpont, N. 1990. Structure and organisation of an Amazonian forest bird community. Ecological Monographs 60: 213-238.
White, G.C. 1998. Program Mark, Version 1.0. [online] Available http://www.cnr.colostate.edu/~gwhite/mark (22 July 1998).
Table 1. Approximate numbers of net-days per year for each of the main net lines operated at Cocha Cashu, Manu National Park, Peru, from 1973-1982. Additional netting took place along several other transects for only a single year.
Table 2. Sample sizes, mean wing chord (mm) and body mass (g) and estimated apparent survival rates (f ) for 14 species with at least 50 individuals captured between 1973 and 1982 in all net lanes at Cocha Cashu, Manu National Park, Peru. Model f (c), p(t) assumes survival rates (f ) are the same for all birds over time, while model f (a), p(t) assumes that birds have different apparent survival rates in their first year of banding (f1) than in subsequent years (f ), possibly because of immature birds or transients. The model selected by AICc is indicated in bold.
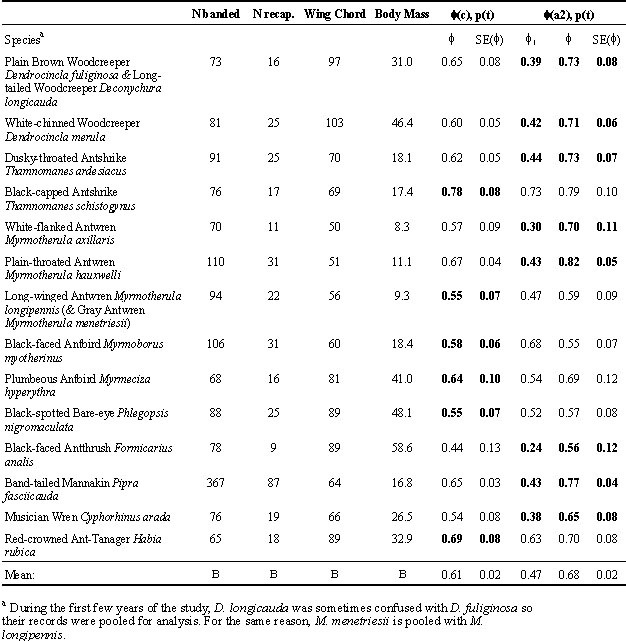
Table 3. Median and maximum movement distances (m) between location of first capture and location of first capture in the next year it was captured, compared with distances between capture locations in subsequent pairs of years. If a species was captured more than once within a year, only the first capture location is included.