S03.5: How Nightingales develop their vocal competence
Dietmar Todt & Henrike Hultsch
Institut fuer Verhaltensbiologie, Freie Universitaet Berlin,Haderslebener Strasse 9, D-12163 Berlin, Germany, fax 49 30 838 5581, e-mail hultsch@zedat.fu-berlin.de / todt@zedat.fu-berlin.de
Todt, D. & Hultsch, H
. 1999. How Nightingales develop their vocal competence. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 193-215. Johannesburg: BirdLife South Africa.Territorial Nightingales Luscinia megarynchos invest a lot of time in singing, and much of this behaviour may consist of elaborated forms of male-male interactions. This communication operates by an exchange of songs and profits from the following mechanisms: a hierarchical memory organisation that obviously supports the retrieval of specific song types, and a song control system that allows individuals to adjust their vocal replies temporally to the start or the end of a neighbour’s song. Song sharing is a consequence of vocal learning that is guided by social factors. Studies on how Nightingales develop their interactional skill have shown that the capacity for vocal matching occurs early in a bird’s life, i.e. before song crystallisation. Addressed time specific responding, like overlapping another bird's songs, however, occurs in the adult singing only and seems to be related to a territorial status.
INTRODUCTION
Avian singing occurs in a fascinating variety of forms. It can be given as a solo-performance or as an orchestration of many different voices (chorus), or it can take place as a display of two particular individuals. In most adult birds, singing is composed of a species-typical pattern that is called ‘song’ (strophe) and that plays a significant role as a unit of interaction. During dyadic encounters, for example, birds can alternate their songs. These alternations in song performance prompt questions of whether and how far they are related to a mutual exchange of messages. In this chapter we will review results of studies that addressed these issues in birds who develop and use a large repertoire of different song-types. These birds have a number of options for responding to each other, thus their interactional accomplishments are particularly interesting.
Interactions by song have been categorised according to formal and functional aspects. From a formal perspective interactions are distinguished, for instance, in terms of relationships that exchanged signals show in the time and the pattern domain (Hultsch & Todt 1982; Wolffgramm & Todt 1982). From a functional perspective, in contrast, interactions are specified in terms of social relationships among signallers and distinguish, for instance, behaviours between partners, such as mates, or behaviours between rivals, such as territorial neighbours. Mated birds often interact via elaborated vocal duets (Todt & Fiebelkorn 1979; Todt et al. 1981; Farabaugh 1982) whereas rivals typically engage in forms of counter-singing, i.e. vocal duels (Todt 1970a,b; Kroodsma 1979; Falls at al. 1982; Kramer & Lemon 1983). Here we will deal with vocal interactions among territorial neighbours and describe how both their time and their pattern specific responses are related to functional aspects.
Birdsong is a learned behaviour, and the study of this learning can help to elucidate how birds develop their interactional skills (reviews in Catchpole & Slater 1995; Kroodsma & Miller 1996). Several characteristics of adult singing develop only if birds have been auditorily exposed to the species song early in life. For instance, if a male has been raised in acoustic isolation, he will fail to develop the normal species typic song patterns. In contrast, with respect to timing of the performance, his vocalisations will be rather normal. That is, he will appropriately segregate his performance into songs of a few seconds duration and silent intervals separating successive songs. Thus, it is expedient to distinguish between the pattern and time domain of singing and examine the impact of experience in either domain on mediating adult competence.
Our model species will be the Common Nightingale Luscinia megarynchos. Males of this species acquire and use a large vocal repertoire, i.e. about 200 discretely different song-types, and extensive field studies have been conducted on both its nocturnal and diurnal singing. Furthermore, we studied the acquisition and development of song in handraised birds, allowing us to explain a number of phenomena obtained from field experimentation.
INTERACTION VIA SONG
Singing is a long distance display and one can expect that oscines birds are well adapted to settle many of their territorial disputes by vocal interactions. For a number of reasons, Nightingales are particularly appropriate subjects for studying this matter: During the day, males of this species behave very much like any other conventional songbird, but they are extraordinary before dawn. With no interference from other species, they sing precisely and with a strong voice, often for several hours and at the same place. In addition, because visual signalling is impaired in full dark, the birds should encode and broadcast messages exclusively in the acoustic mode. This assumption was tested by playback experiments during the nocturnal singing of Nightingales: We found that birds did not or only rarely approach loudspeakers, and this behaviour allows us to address rules of both time- and pattern-specific responses to conspecific song (Fig. 1).
Time specific responses
Many animals avoid simultaneous production of vocal signals, and by examining how temporal adjustment is distributed in the field, researchers uncovered a system of temporal interaction strategies. In songbirds this avoidance behaviour can be observed as segregation of performance times on a larger scale, including heterospecific avoidance, and as an alternation of songs on a finer scale. A basic functional aspect of signal alternation and other time related behaviours is the use of temporal signal displacement as a strategy to prevent a bird’s own vocalisation from being masked acoustically (Cody & Brown, 1969; Ficken et al. 1974; Wassermann, 1977; Gochfeld 1978; Hultsch & Todt 1982). Temporal adjustment is especially clear during the nocturnal singing of Nightingales, and can be specified as follows.
In a population of territorial birds, individuals exist who - during dyadic singing - preferentially start their songs shortly after a neighbour has terminated his preceding song. These responders are called ‘inserters’ or ‘alternators’, and are distinguished from other responders called ‘overlappers’. Overlappers preferentially start their songs some time before a neighbour has finished a preceding song. However, besides inserters and overlappers one can find individuals who obviously do not adjust their songs to a neighbour's vocalisation, but rather seem to follow their temporal self-program. These birds are called ‘autonomous songsters’ (Hultsch & Todt 1982). There is evidence that the three distinguished behaviours cannot simply be explained as individual diversities, but rather highlight three different interaction roles. To apply the role concept here is appropriate for two reasons. First, each ‘role’ reflects a specific strategy that is used in relation to season and social context, and second, depending on these variables individuals may change their ‘role’.
Functional aspects
The three roles serve different functions. Inserting is a very common strategy which obviously serves acoustic avoidance, mutual listening, signal detection and responding. Overlapping, in contrast, which is predominantly found at the beginning of the territorial season, allows a bird to challenge and even to repel a competitor, mainly by jamming the other bird's signals. Autonomous singing, finally, occurs especially in territorially well established males and thus seems to signal a dominant status.
Role changes
Changes of interaction roles occur in various forms. For instance, a male acting as an overlapper towards a neighbour X may behave as an inserter when interacting with another neighbour Y. Or, if an autonomous songster is exposed to playbacks simulating an intruder to his territory, he may switch to time specific responding. More commonly, and regardless which role an individual had taken during night-time, males switch to autonomous singing when the daily dawn chorus begins. Concurrently, they reduce the duration of their silent intersong intervals. Such changes can be viewed as a correlate of both state modifications and changed environmental conditions, e g. the often extremely noisy background of chorusing.
The significance of temporal interaction strategies stimulated us to examine the mechanisms by which temporal adjustment is controlled. It turned out that song onset latencies were rather precisely tuned to the song start of the other male in the overlapper, and to the song end of the other male in the inserters. In both the overlappers and inserters, latencies peaked at about 1s, showing that birds are indeed listening and responding to each other (Hultsch & Todt 1982). This notion was substantiated by an interactive playback experiment that examined the inserter role in more detail. In the experiment, both the latency with which stimulus songs were broadcast after each song utterance of the test bird and the duration of the stimulus songs were varied. The results, illustrated in Fig. 2, revealed an impressive flexibility in the temporal adjustment of the test bird. Furthermore, there was a striking after-effect following the playback trials: the normal timing of song delivery was not immediately resumed. Rather we recorded prolonged intersong intervals for a couple of songs, before these gradually approached normal duration (Fig. 3). Such behaviour, in particular, suggests that the bird ‘waited’ for another song to occur. In line with results from other playback experiments, simple stimulus/response mechanisms that would account for the adjustment of song onset times cannot explain these findings. Rather, they point to a basic motivational variable like being in an ‘interactive state’ that may underlie the diverse facets of temporal interactions.
In functional terms, some aspects of the documented performance roles remain unclear. One would expect, for example, that the overlapper as well as the addressee should suffer from signal masking. In fact, the consequences of overlapping are asymmetrical, for two reasons. First, the overlapped bird often interrupts his songs or may even terminate singing. Second, the overlapper often considerably prolongs his own songs, thus leaving large parts of them undisturbed. Thus, acoustic jamming of another bird’s signals may not be the prime function of the overlapper’s behaviour. Rather, increasing evidence suggests that a distinct message is encoded in overlapping, namely a vocal threat or challenge (Todt 1981; Hultsch & Todt 1982; McGregor 1991; Todt & Hultsch 1996). This issue prompted questions on whether a given interaction role might provide cues that can be decoded by ‘third party’ listeners. Relevant evidence for such possibility came recently from a study of Naguib and Todt (1997) who showed that territorial Nightingales that were exposed to simulated dyadic vocal interactions indeed responded more strongly to a simulated overlapper than to a simulated inserter.
In summary, analyses of time specific relationships between exchanged vocal signals reveal that songsters do not just avoid temporal signal overlap, but show differentiated performance strategies. This behaviour highlights the question of how birds respond to the specific pattern of one another’s song signals.
Pattern specific responses
During auditory exposure to a singing conspecific, territorial birds may modify the patterning of their song performance. Such modification can affect different hierarchy levels of song organisation, and at the same time also reflect a tonic or a phasic response. In a tonic response, a bird alters the quality of his singing gradually over an extended span of time, for instance by increasing the volume or duration of his songs. In a phasic response, in contrast, a bird selects a particular song type of his own repertoire and vocalises this song as a reply to a stimulus song. Examples of this form of responding are song matching and cases of co-ordinated switches between bouts of songs. Song matching is quite common across species (Lemon, 1968; Smith & Reid, 1979; Falls et al. 1982; 1988, Beecher et al. 1996), and is particular impressive in songsters who, such as Eurasian Blackbirds (Todt 1970a, 1975, 1981) or Nightingales (Todt 1970b, 1971a; Hultsch 1980), have large song repertoires and sing with an immediate versatility (e.g. A-B-C-D-..). Coordinated song type switching, in contrast, is found in species that repeat renditions of a particular song type several times before starting to sing another one (e.g. A-A-A-B-...), like e. g. Abessinian Ground Thrushes (Todt 1971b), Great Tits (Falls et al. 1982, Mc Gregor et al. 1992), Western Meadowlarks (Falls & d’Agincourt 1982; Horn & Falls 1988) and Song Sparrows (Kramer & Lemon 1983, Kramer et al. 1985; Nielsen & Vehrencamp 1995).
Pattern specific responses depend on species typical properties, but they also build on prerequisites in the vocal repertoires of the individuals involved. This dependence is particularly evident in song matching. Per definition, song matching (synonymous term: ‘equivalent response’) states that a song X is responded to by a song with an equivalent pattern X’. Thus matching can take place only between individuals who share song-types in their vocal repertoires. If, however, individuals interact with convalent responses (synonymous term: ‘vocal supplementing’), a stimulus song X is responded to by a song Y that can be regarded as a sequential continuation of ‘X’. Thus for interactions with convalent responses neighbours do not only have to share parts of their repertoire, but also must have corresponding sequential associations among song-types. In neighbouring Nightingales, such sharing of ‘singing programmes’ may have an interesting outcome: A convalent response would allow a male to become a ‘sequential leader’ in a vocal interaction (Todt 1971b; Todt & Hultsch 1980).
Given the remarkable differences that species show in various domains of their singing it is not surprising that several hypotheses have been proposed to explain the proximate functions of pattern specific responses in vocal interactions (Todt 1971a, 1975, 1981; Kroodsma, 1979; Falls et al., 1982; Krebs et al.,1978, Kramer & Lemon, 1983; Hultsch & Todt, 1986). For vocal matching, which to date is the most extensively studied response category, such hypotheses range from very straightforward ones, like ‘addressing an opponent’ (Todt 1975) or ‘sending a keep-out signal’ (Falls 1985; Shackleton & Ratcliffe 1994), to more controversial ones like ‘distance estimation’ (Falls et al. 1982; Morton 1982; Naguib 1997) or ‘attracting the attention of a third party’, like females or other males (Todt 1981).
The basic ‘addressing’ account of vocal matching is a candidate mechanism to deal with the general problem of signalling to a particular male in a community of songsters. The interesting issue is that a more specific message of the signal can be encoded in its timing, as has been shown in Nightingales and Eurasian Blackbirds (Wolffgramm & Todt 1982; Hultsch & Todt, 1986). These species use song matching in two temporally different forms. In the rapid matching, a song-type A is responded to by A’ quickly after its onset. Thus the matcher overlaps the other male’s song. In the delayed matching, in contrast, the matcher waits until the other bird has ended his rendition of A, thus inserting the response into the intersong interval of that male. The message to infer from rapid matching is similar to that one described for temporal overlapping, a vocal threat. Rapid matching dominates at the beginning of the singing season when songsters set up territories, i.e. mainly during highly agonistic interactions. In contrast, delayed matching can be found particularly when territories are established and when other song features of neighbours indicate that they sing in a somehow relaxed way. This response, therefore, was interpreted as a vocal ‘greeting ritual’ (Todt 1974, 1981).
Viewed from a functional perspective of interaction processes, it is important to save the salience of a signal and hence to keep its occurrence at an optimal level. This view explains why vocal interactions, like many other kinds of signal exchange, are not perpetuated during the entire time two neighbours are singing in parallel, but rather occur sporadically. In proximate terms, in contrast, the length of interactional phases seems to be limited by properties of a bird’s endogenous program of song delivery (self-program), which affect the access or retrieval of a particular vocal pattern in response to a heard song. If, for example, a bird has developed strong sequential associations among song-types (i.e. treats these as a super-unit), he may not be able to abandon that train of songs and give a matching response. Male Eurasian Blackbirds and Nightingales develop several ‘subrepertoires’ of sequentially associated song-types and playback experiments suggest that such organisation indeed restrains the options to retrieve a particular song-type (Todt 1971a; 1975; Hultsch 1985). If males, in contrast, share such ‘subrepertoires’, vocal interaction can be facilitated. Correspondences here have implications not only for the probability and speed of responses, but also for the response category (equivalent or convalent) that a male is going to choose.
In summary, analyses of time and pattern specific relationships between exchanged songs reveal that birds perform differentiated forms of vocal interaction. Vocal interaction builds on certain prerequisites, like sharing of signal repertoires if matching is to occur. For other vocal responses the prerequisites can be more specific, and often they are also more difficult to assess by the experimentor. Convalent responses, for instance, require fixed sequential associations among particular types of song, and mutual sharing of such singing programs can allow a variety of specific vocal interactions. It is expedient to assume that vocal learning plays a crucial role in establishing such prerequisites. We discuss this issue by asking how Nightingales acquire their vocal competence and how this learning provides them with a sharing of both song repertoires and song performance programmes.
ACQUISITION OF SONG
In conjunction with the evolution of song as a culturally transmitted behaviour, birds have developed a system of adaptations which ensure the biologically adequate use of song as a signal system. Among them are phenomena like age-bound auditory learning (sensitive phases), a preference for the species-typical song pattern (signal selectivity), and a preference to acquire song from a socially significant individual (social selectivity). Large differences among species exist among species in the extent to which these mechanisms become effective during the perceptual phase of song acquisition, and constraints seems to be tightly related to the particular ecology of a given species or even population (for reviews see: Kroodsma & Miller 1982; Slater 1989; Marler, 1987, 1991; Baptista 1996; Beecher 1996; Kroodsma 1996).
Repertoire acquisition in Nightingales
In its singing, an adult Nightingale may use more then 200 different types of songs. These song-types are discretely different and are reproduced as stereotyped pattern compounds. In addition, males often share song-types as rather exact replicas, and repertoire congruency of neighbouring birds may be up to 75% (Hultsch 1980; Hultsch & Todt 1981). Thus Nightingales clearly do not create their vocal diversity by continuously improvising on a basic pattern or theme. Rather, vocal learning by imitation is the candidate mechanism to account for repertoire development in this species. The large repertoire prompts questions on whether it is acquired during a rigidly defined time window that would close early in ontogeny, e.g. before dispersal, here fall migration. The experimental evidence obtained on the timing of song learning in Nightingales is not surprising: As with other oscine birds, Nightingales will begin to acquire songs around fledging (ca day 15 post hatching). But, setting aside cyclic or seasonally induced bias on learning, males were found to acquire new song-types until their third year of life in the laboratory (Wistel-Wosniak & Hultsch 1992).
Nightingales learn in a socially selective manner. As tested in choice experiments (Todt et al. 1979), acquisition from taped exposure to master songs alone turns out to be extremely poor, whereas males readily acquire these songs provided a social tutor (the human caregiver) is present during the playback. Whether or not the birds ‘accept’ the human tutor is crucially affected by the age when handraising begins: only if handraising begins before day 8 post hatching, are the biological properties of the tutor insignificant. Thus, interactions during handraising may induce imprinting-like processes that prepare a young bird to learn his songs from the human caretaker. Nevertheless, social selectivity of Nightingales decreases as they become older. For instance, males will acquire new songs heard from tape alone at an age of about 9 months (Hultsch 1991a), which roughly corresponds to the time when free ranging birds may be further exposed to conspecific songs in their winter quarters (Nicolai, 1976). The decline in social selectivity turned out to be not a mere consequence of age, but rather seems to be bound to previous experience of conspecific songs in a social context (Todt et al. 1979; Todt & Böhner, 1994). This finding suggests that one function of the social tutor could be to somehow calibrate the acquisition system by providing information about the adequate species-typic song organisation. These features may then serve as a basis against which further experience is compared. Thus song acquisition may emancipate from the social model later in life. Such mechanism seems adaptive in species where males acquire large repertoires of song-types, and Nightingales are clearly disposed to do so.
To teach young Nightingales, we expose them to a number of master songs that in our standard design are spaced by normal intersong intervals (duration ca. 4 s.) and played back through a loudspeaker while their tutor is present. The songs to which the subjects are exposed are normal, species typical song-types drawn from performances of free ranging males. However, only song-types that had been found to (a) be acquired by the subjects with equal facility and (b) not occur in a bird's repertoire if he had not heard them during an appropriate teaching session, are included in our catalogue of master song-types. To compose a learning program, particular songs are selected at random from this catalogue and recorded on tape to form a particular string of master songs. In the standard design, each song in a string is a different song-type and, likewise, each of the different strings to which a subject is exposed during the period of tutoring consists of a unique set of song-types. This means that song-types occurring in one string do not occur in another. In addition, birds do not have auditory access to any of the master song-types outside of a specific set of training sessions (Hultsch et al. 1984). By using particular sets of learning stimuli in a particular learning situation or tutoring regime only, we label such situation or regime, and this procedure allows inferences on whether and how a particular exposure variable has influenced the singing of the tutored subjects. Additional checks for an impact of variables are done by an analysis of audiovisual recordings which permit access to, for example, an individual bird's motility during a given tutoring experiment (Müller-Bröse & Todt 1991).
The first step in the analysis of the developed song patterns consists of checking for imitations of master song-types that were presented during the tutoring. Normally the structural details of models are quite faithfully copied and deviations concern mainly additions or deletions of some syllables in the first song sections. Hence, the identification of imitations is straightforward and, as a rule, unambiguous in Nightingales (Hultsch & Todt 1989b). In a second step of analysis, we assess the birds’ learning success, i. e. the proportion of imitated song-types to master song-types. Finally, variables that may have affected the learning are identified by comparing the acquired song-types and the mode of their performance. The study of organismal variables, such as age and social experience, is a prerequisite to investigating the experimental variables which comprise, for example, features of the
tutoring regime such as exposure frequency, the number of master songs in a tutored string, presentation rate, serial position, and the distribution of tutoring sessions in time.Although song acquisition clearly depends on experimental variables, it can be remarkably resilient to changes in such variables. For example, while the acquisition success of Nightingales is low (app. 30%) for songs experienced only 5 times, birds imitate around 75% of those song-types which they heard 15 times. A more frequent exposure does not significantly improve acquisition success (cf. Fig. 4). Also, the number of songs in a string can be considerably increased (e.g. from 20 to 60 song-types) without raising exposure frequencies accordingly. As the birds cope well with such an increase in the number of stimuli to be acquired (Hultsch & Todt 1989a), the results contrast with paradigms from learning theory, which state that exposure frequency must grow proportionally with the number of stimuli to be acquired (review in Crowder 1976). Implications of our findings are relevant to the issue of song acquisition as a special process or template learning (Marler 1976) and any inquiry into the memory mechanisms has to take into account that specific adaptations are involved in the process. The functional properties of acquisition mechanisms can be traced by a broad array of experiments that include the perceptual and motor phase of song development. In the following we will exemplify representative findings from our behavioural approach.
Sequential learning in Nightingales
The large repertoire of distinct song-types in Nightingales allows to study the impact of learning on different levels of song organisation. In the following we describe how learning affects levels beyond that of a song, and how it can shape the performance of long sequences of songs (inter-song level). We have investigated this issue by exposing Nightingales to an increased number of master songs. During the tutoring, these songs were presented as units of several different master strings, each of which the birds experienced 20 times. When song repertoires performed by the trained birds were examined for rules of song-type sequencing, we found a surprising effect:
The Nightingales' singing clearly reflected information about the context of master song presentation. In other words, males produced imitations acquired from the same master string as sequentially associated song-types and segregated from imitations acquired from the other strings. This achievement was described by the term context effect (Hultsch & Todt 1989c), and explained as follows. Nightingales acquire and memorise a given string of master songs as a kind of super-unit which, upon retrieval, they separate from another super-unit, i.e. from songs learned from another master string. These units are termed context groups. The size of such groups is determined by the length of a tutored string, or the number of different master songs that a bird had experienced in the same temporal context. Further experimentation allowed us to characterise the time between exposures to different master strings that was necessary to generate a context effect. If strings were separated by less than 5 minutes, no clear context effect could be assessed (Hultsch & Todt 1992a).
The formation of context groups turned out to be a basic feature of song learning in our birds. However, detailed inquiries into the song sequencing of context groups revealed another significant effect. Within performance episodes of context groups, we detected smaller subsets of sequentially associated song-types, which we called 'packages' (Hultsch & Todt 1989b). In functional terms, package formation describes the fact that a larger body of serial data (i.e. information from a string of master songs) is segmented into subsets of sequentially associated items. Such associations have a number of characteristics:
First, the packages describe song-type associations which are hierarchically inferior to the context groups. Consequently, one can sketch a structural hierarchy that in a top-down order reads as follows: First, context groups are composed of song-type packages and packages are composed of songs. Second, each package holds a limited number of song-types only, and frequency distributions of package sizes reveal a prominent peak between three and five types of songs. Third, within a package, sequential associations among song-types are stronger than associations between different packages. Nevertheless, song-type sequencing within a package can deviate from a unidirectional succession (schema: A-B-C-D-E) and show more flexible permutation modes (e. g.: A-B-A-E-D-E-). Such complex combinatorial relationships among package members indeed substantiate the notion that package groups establish a level of hierarchy in the representation of song-types. Finally, different birds exposed to the same master string usually form different packages or show different boundaries between packages, respectively. Therefore, package groups can be characterised as self-induced associations.
As for the mechanisms underlying package formation, a process was postulated that segmented the information about a succession of master songs, which had been temporally coherent during exposure. This raised the issue of when after exposure and where in the subsystems mediating between auditory exposure and vocal production of songs such a segmentation would take place. Theoretically, it could be part of the acquisition, the storage, or the retrieval system (Hultsch & Todt 1989b).
To date there is substantial evidence suggesting that package formation results from a process which segments serially presented information during auditory acquisition. One study used an indirect approach and tested Nightingales raised in acoustic isolation. The examination of whether song-types packaging would occur in these birds yielded negative evidence (Wistel-Wozniak & Hultsch 1992). In a more experimentally oriented approach (Hultsch 1992), Nightingales were exposed to strings of master song-types which instead of normal intersong intervals (4 s) contained either prolonged (10 s) or reduced interval duration (1 s). If the birds singing showed that the size of developed packages was not affected by these treatments, this would favour the hypothesis that package formation is a retrieval based phenomenon. The results allowed us to reject this hypothesis. Rather, the study provided evidence that package formation is a correlate of auditory learning and affected by two properties of the acquisition mechanism: a capacity constraint memory buffer and a time window. Additional support for package formation as an early achievement of song learning came from a third line of evidence. The package effect gets masked when the serial order of performed song-types becomes more stereotyped. This masking could be induced, for instance, when the exposure frequency to stereotyped master strings during acquisition was high, e.g. 100 times. ('serial order effect' , Hultsch & Todt 1992b).
In summary, our studies on Nightingales have shown that birds acquire information encoded in the serial succession of master song-types presented during their training. Concurrently, the birds develop additional hierarchy levels such as packages and context groups. In contrast to the self-induced package groups, both serial song order and context groups do clearly reflect the organisation of input during auditory learning and consequently are distinguished as exposure-induced compounds.
Cognitive accomplishments in song acquisition
The discovery of the context effect stimulated us to ask whether song learning would entail cognitive processing, such as a cue-related categorisation of songs. Recall that handraised Nightingales produce sequentially associated and segregated songs according to the program from which from songs were learned. The characteristics of context group formation suggested that birds use a time based cue to segregate these groups during auditory acquisition. However, it remained to be determined, if context group formation reflects a process of categorisation, such as sorting songs as ‘members’ of string A, B, or C, etc. To deal with this issue experimentally, we introduced specific contextual variables to our learning programs that might act as cues for categorisation.
We pursued two approaches. In one approach, we controlled for the mere temporal account of the context effect by adding different cues to the songs of one and the same master string. Thus, we sought to abolish the integrity of a learning context by using different cue labels, and asked whether subjects would parse the string according to these cues (see below under 'splitting a learning context'). In the other approach, we examined categorisation by adding same cues to different master strings. Thus, we sought to combine different learning contexts through shared cue labels, and asked whether subjects would treat these contexts as a super-unit according to the associated cue (see below under 'lumping learning contexts').
Splitting a learning context
Contextual cues were provided by the simultaneous presence of two human tutors during the training, who switched roles during the playback of an otherwise coherent master string. The tutors, one familiar and one unfamiliar to the birds, were positioned with their speakers at opposite corners of the bird room and, while the playback was running, the one or the other speaker was activated (Fig. 5).
The analysis revealed that birds had acquired more songs from the unfamiliar tutor and also preferred to sing them over those acquired from the familiar one. This finding suggests that the social reference of the songs presumably played a role here. The singing patterns of two males was indicative of context-based group formation. These males associated imitations sequentially according to the tutor who had presented them. Thus, in addition to demonstrating song discrimination, they had also parsed and memorised songs in relation to social cues.
Lumping learning contexts
In this experiment, contextual cues were provided by the presence or absence, respectively, of a conspicuous visual stimulus (strobe light) that was paired with the acoustic stimuli. Nightingales were exposed to nine master strings that they would normally memorise as nine different context groups. Four of these strings were presented in the strobe regime, whereas in the remainder strings no such stimuli were used. The temporal succession of strings during the tutoring period was randomised.
The birds’ singing revealed that these learning programs had affected both the acquisition and the performance mode of acquired songs. For instance, males acquired more songs from the strobe regime than from the control regime, suggesting that exposure to the strobe induced an increased attention or arousal in subjects. Two other effects were relevant to the issues raised above: (1) Males preferred to sing strobe imitations over those experienced through hearing alone. (2) We could identify sequential clusters of songs that were made up of imitations from either the strobe regime or the control regime, even when the respective models songs had been members of different master strings during the tutoring. Because none of these effects could be accounted for regime related differences in the acoustic stimuli, we conclude that the birds encoded and stored master songs according to regime category.
Referring to our current model on the hierarchical representation of acquired information, these findings can be accommodated by postulating some kind of 'top level hierarch' that links song material acquired from different master strings according to tutoring context. The fact that this process was induced by the simple stimuli as used here, documents that the song acquisition system of birds is extremely sensitive or 'predisposed' for the processing of non-acoustic contextual information that is associated with the acoustic experience. From all that is known about the functional significance of singing behaviours, such an adaptation makes strong sense. Therefore it is now expedient to examine in more detail the impact of cues that are biologically more relevant than the ones tested here. As suggested by the former experiment, candidates would be, for instance, categories as 'song-types heard from a particular individual', 'song-types heard at a particular location', or 'song-types specified by a particular quality'. Functionally, an association and cue related categorisation of acquired song material would allow the bird to adapt repertoire delivery to its individual acoustic and social environment.
Field data supporting the view that associative learning may play an important role in song acquisition have been obtained from the Thrush Nightingale Luscinia luscinia (Mundry in prep), a twin species of the Common Nightingale. In areas where these species are sympatric, genetic Thrush Nightingales are found who sing songs of either species. According to phonetic and syntactic characteristics, their different songs could be sorted into three classes: Nightingale, Thrush Nightingale, and songs with mixed characteristics. These data suggest that the males learned from two species of tutors. In addition, their performance was clearly not random with respect to the sequencing of these pattern classes. Rather, males performed them sequentially lumped in coherent bouts. Thus, temporal and social segregation of exposure contexts were reflected in the singing program of the adult birds.
DEVELOPMENT OF SINGING
The ontogenetic development of singing in Nightingales shows a number of traits that are wide-spread across oscine birds. For example, the early phase of auditory learning is temporally segregated from the phase of vocal production by several weeks. Vocal activity of young birds may cover a time span that often lasts for several months until they reach adult vocal competence (Fig. 6). Early in life, birds perform temporally coherent arrays of vocalisations that first are phonetically amorphous and only gradually improve in structure. Referring to the profile of developmental progress, Marler and Peters (1982a) have suggested a tripartite model which distinguishes among (a) subsong, (b) several stages of plastic song and (c) crystallised fullsong. A detailed study of these stages can uncover rules that describe the 'ontogenetic trajectories' of singing, i. e. transitions from incomplete song precursors to their crystallised expressions.
Ontogenetic trajectories
In birds that, like Nightingales, develop large repertoires of songs, ontogenetic trajectories are found on different hierarchy levels, for instance, within songs or between them. At the within-song level one can distinguish the following traits. During early stages of ontogeny, birds often sing incomplete songs; i.e., some song constituents may be missing. In addition, the serial succession of song sections may be inverted, which may result, for instance, in the final trill-section of a song being produced ahead of the normally preceding note complex. Thus, the syntax of songs is not initially stereotyped. At the same time, however, the phonetic morphology of patterns is sufficiently elaborated to allow for an easy identification of song-type precursors. In other words, pattern phonetics takes its adult form ahead of syntax, which 'crystallizes' only at about 10 months of age. Interestingly, the phonetic or syntactic quality of imitations that emerge relatively late in ontogeny is not inferior to the quality of imitations produced at an earlier age. This finding suggests a developmental trajectory which does not build on vocal 'experience' with a particular output, but concerns a general progression in motor competence or skill (Hultsch 1991c).
Besides involving trajectories concerning the pattern structure of vocalizations, ontogenetic progression of singing also proceeds in the time domain of singing. Because both the duration of vocal compounds and their temporal segregation must be shaped, trajectories in the time domain are highly interrelated and follow complex rules. The adult time structure of singing (songs alternating with silent intervals of about the same duration; see Fig.1) is the last performance feature to crystallise; its adult form is achieved only at an age of about 11 months (Kopp 1996).
On a higher level of song organisation, the following rules can be characterised. Imitations which, in the adult performance, are identified as members of the same package emerge, quite consistently, together in time. In addition, throughout ontogeny these precursors are sequentially associated in the same way as in the adult singing (Hultsch 1989). The association of different packages, i.e. the development of context groups, in contrast, seems 'delayed'. However, the time structure of singing gives some indication that these groups are significant building blocks of the performance already during ontogeny. During the phase of continuous vocal production, for example, intervals between imitations acquired from the same master string or context group were significantly shorter than those intervals assessed when the birds switched to imitations of another context group (Hultsch 1993). This effect suggests that access to the stored representation of song-types may be quick or delayed, depending on whether retrieval is from within a given context group or from a different context group. Alternatively, the differences in intervals could reflect decision times for retrieving patterns acquired from same or different contexts. In conclusion, on the inter-song level, the ontogeny of song material indeed reflects properties of the song-type association groups referred to earlier. Trajectories do not only substantiate the view that these groups are memorised and encoded as higher levels units of song organisation, but also suggest that retrieval follows hierarchical mechanisms of action selection.
Repertoire development in Nightingales
When birds enter into the final stage of song development, i. e. reach their adult form of singing, they often omit a certain portion of imitations from their repertoire (Marler & Peters 1982b). In Nightingales, however, this constriction is much less marked than in other species (Hultsch 1991b). Instead, an increase in repertoire size along with age is more common in this species (Freyschmidt et al. 1984; Wistel-Wozniak & Hultsch 1993). Such increase results either from additional song acquisition or from the development of new songs. The latter mode of repertoire enlargement, that enhances the vocal individuality of a songster, is achieved by the development of novel inventions or new recombinations of songs.
Nightingales may develop song-types that do not contain material from the learning experiments, and so are completely new. Both during ontogeny and adult singing, genuine inventions occur as coherent subsets in the singing, which results in an alternation of performance phases containing acquired imitations or novel inventions (Hultsch et al. 1998). Males classified as poor learners develop a larger proportion of inventions than those classified as good learners. Thus one may speculate whether, at least in handraised Nightingales, song invention reflects a predisposition to develop a vocal repertoire of a certain size. A similar effect can result when Nightingales generate individual specific song-types by recombining parts of imitated songs in a novel way. Interestingly, such recombinations are limited to material of song-types associated within the same package group (Hultsch 1990). Packages also remain closed to additional song-type acquisition. Context groups, in contrast, can be enlarged by newly acquired songs. When, for example, birds are exposed to a master string with two parts, a familiar portion heard during the early tutoring and a novel portion containing new song-types, the novel portion is integrated into the same context group as the familiar song-type sequence (Hultsch 1991c).
The sustained ability to acquire novel songs during vocal ontogeny is in line with our observations of trained birds who, instead of being housed in isolation, were housed together. Allowing them to interact with each other vocally shaped the composition of song repertoires, and the performance frequency of shared song-types also clearly converged. Thus, additional learning coupled with a shaping of the performance towards convergence can lead to a sharing of at least parts of repertoires among conspecific neighbours.
Development of singing interactions in Nightingales
Nightingales housed together were often found to sing parallel in time during the phase of plastic song. Although they may produce similar vocal patterns during this phase, their performances usually did not exhibit the clear pattern specificity of interactions that occurs in adult males. Nevertheless, the evidence suggests that the capacity for vocal matching may emerge early in a bird’s life. When subjects reached the phase of crystallised song, the amount of ‘parallel singing’ decreased, and after developing the species-typic silent inter-song intervals, birds preferentially alternated their songs.
To elucidate further how Nightingales develop their time and pattern specific responses to conspecific song, the above observations were supplemented by a more standardised approach. We exposed birds, who after initial tutoring had been housed in isolation, to two additional playback experiments during the phase of plastic singing. The experiments were segregated in time by four weeks (March and April). Song strings used in each playback contained 12 different song-types with 6 of them familiar and 6 new to the birds. In the early experiment, birds tended to interrupt their singing immediately after the onset of stimulation, but resumed singing when the stimulation stopped. No clear evidence could be obtained for pattern specific responses. Although this outcome seemed different from observations under communal housing conditions, it was not surprising. First, our experiment simulated exposure to the singing of an adult conspecific. Second, the playback lasted for a short time only (4 trials, each 5 min), and thus did not allow the birds to become ‘familiar’ with it, a variable that often promotes pattern specific responses (Todt 1970b).
During the second experiment, conducted later in age, our birds responded more specifically. First, they modified the temporal patterning of their vocalisations and produced a kind of pattern alternation, although they were still in the phase of continuous singing. That is, they inserted their own songs into the silent intervals between master songs, and such temporal adjustment was remarkably skilful during the playback of familiar master songs. After a playback trial, however, birds resumed their former continuous style of singing. Second, familiar master songs elicited pattern specific responses, several of which matched a given master song. Interestingly, birds tended to suspend singing when the stimulation began to play the novel master songs. There was no evidence of long-term after effects induced by the experiment. That is, neither was the progression of vocal development accelerated, nor was the onset of song crystallisation faster in the test birds. Finally, vocal replies that, such as song overlapping, encode a clear message, were only given after birds had reached the final phase of adult song, and were restricted to those individuals who were allowed to attain a territorial status.
In summary, as with the ontogeny of singing in general, the ability to interact vocally is involved in the development of motor competence. Practice seems to play a particular role in producing the regular alternation of silencing and activating the vocal tract in response to stimulus songs. Pattern specific responding, in contrast, builds on perceptual accomplishments that are already crucial to the early auditory acquisition, like auditory discrimination of song patterns. Both the ‘listening’ and the behaviours analogous to vocal matching point to a sustained significance, and presumably functional elaboration of, the perceptual system during vocal development.
CONCLUSION & PERSPECTIVES
In the previous sections we have described various facets of how Nightingales acquire, develop and use their singing. To complete the picture, we will now address the functional properties of intrinsic mechanisms that mediate their vocal competence.
The singing behaviour of Nightingales reflects a structural hierarchy that, in a bottom-up order, is given by the units that compose a song (intrasong level), by songs, and finally, by sequences that are composed of songs (intersong level). Inquiries into the role that individual experience plays in the implementation of pattern repertoires and repertoire organisation were particularly productive when carried out on the intersong level. Thus, acquired song-type associations not only reveal a hierarchical representation of the memorised information, but their characteristics also allows to examine the mechanisms underlying the auditory or vocal acquisition during song development.
Acquisition
Song acquisition of Nightingales has been explained as a co-ordinated operation of three mechanisms: a 'short term memory', a 'recognition memory' and a battery of 'submemories' (Hultsch & Todt 1989c). Properties of the short term memory cause a segmentation of serially coherent master strings into different packages of information. Evidence exists that this segmentation results from two constraints, a limited capacity and a time constrained memory span (Hultsch 1992). Properties of the recognition memory, in contrast, identify stimulus patterns as novel or familiar and categorize information of the familiar patterns by song-type, package and context group. Acquired song material is then further processed in a battery of submemories, each of which stores information about a given string segment (a package). Parallel data processing in a battery of submemories would explain why long master strings are learned as effectively as short ones, even when heard only 10 to 20 times. However, since each submemory is supposed to hold information from a given string segment only, an additional process has to be postulated that somehow associates those packages that were developed from a given context group. The proposed acquisition system predicts that the first exposure to a master string would play a key role in the acquisition of serial information on song-type sequencing. Recently, this prediction was confirmed by experiments in which the serial succession of song-types in a master string was altered upon subsequent exposures during the tutoring (Hultsch & Todt 1996).
Representation
Rules of song retrieval reveal that the representation of memorised song material is organised hierarchically. Some of these rules can be identified even during vocal development. Thus, ontogenetic trajectories of the three hierarchy levels which, in a top down order, were described as context groups, packages and song-types, develop in a way that allows us to detect their particular features early on. To recall one example, the temporal diversity of intervals within and between context groups (shorter duration within than upon switches between context groups) presumably points to properties of intrinsic pattern choice or retrieval. The effect could imply that access to the stored representation of song-types is quick or delayed depending on whether a retrieval 'program' from a given context group is already active (i.e. a non-switch) or not (i.e. a switch; Hultsch, 1993).
Retrieval & performance
The way birds perform context groups is particularly useful for exemplifying the complexity of retrieval mechanisms. Performance appears to be regulated by constraints acting on both broad and a narrow scales. On the broad scale, i.e. along with consecutive episodes of singing (‘bouts’), context groups show a balanced sequential distribution. Take, for example, a bird that has acquired songs from five different context groups (A, B, C, D, E). If he frequently performed songs belonging to group A, C, and E in the first bout , he will prefer the songs from group B and D in the second bout. On a narrow scale, the performance of a context group is limited in length to appr. 8-15 song renditions, irrespective of the size of the group (e.g. 3, 15 or 30 song-types). At transitions between songs from two context groups, song renditions appear interdigitated. That is, as songs from group A would ‘fade out’, those from group C would ‘slip in’. To examine these effects further, we are using a model that operates by a system of parallel retrieval buffers. In addition, we investigate questions of whether and how intrinsic decisions take place in a hierarchical top-down order, i. e. from context groups to the packages and finally song-types.
In adult birds, a hierarchically prestructured repertoire could be a mechanism to facilitate retrieval in situations demanding rapid vocal responses, e.g. during interactive countersinging. Here, centrally or auditorily mediated decisions on 'what to sing next' would not have to be made among the entire pool of developed song-types. Rather, decision steps and decision time would both be reduced by using a search routine that subsequently addresses a particular subset of patterns only.
This view has implications for modelling approaches to the control of singing behaviours. In the past, these models referred to the ‘song’ as the unit of decision and retrieval. In addition, although assumed to be relevant, memory was not a functional component in the early approaches (Hinde 1958; Todt 1968, 1970a, 1975, 1977; Thimm et al. 1974; Todt & Wolffgramm 1975; Dobson & Lemon 1977; Slater 1978, 1983; Whitney 1981). Our evidence on acquired song assemblies, a potentially widespread phenomenon among oscine birds, calls for a revision of the classical control models of song performance. New paradigms will not only have to account for hierarchical representation of song material, and consequently top-down or bottom-up mechanisms of decision making. They will also have to allow for parallel computation and decision-making on different hierarchy levels and include more elementary pattern units. For heuristic reasons, both the singing performances and the characteristics of vocal responses obtained from experimentally tutored birds will be invaluable here.
ACKNOWLEDGEMENTS
Many people provided priceless help in getting and handraising our birds, doing experiments and data analyses. Among them are Petra Ambrugger, Nicole Geberzahn, Marina Hoffmann, Roger Mundry, Friederike Schleuß, Lars Schrader, Gisela Schwartz-Mittelstädt, Marc Naguib and Alexandra Wistel-Wozniak. We also are also most grateful to Irene Pepperberg for comments on the manuscript. Our work was supported by the DFG and the BBAW.
REFERENCES
Baptista, L.F. 1996. Nature and its nurturing in avian vocal development. In: Kroodsma, D.E. & Miller, E.H. (eds). Ecology and evolution of communication. Ithaka, NY; Cornell University Press: 39-60.
Beecher, M.D. 1996. Birdsong learning in the laboratory and field. In: Kroodsma, D.E. & Miller, E.H. (eds). Ecology and evolution of communication. Ithaka, NY; Cornell University Press: 61-78.
Beecher, M.D., Stoddard, P.K., Campbell, S.E. & Horning, C.L. 1996. Repertoire matching between neighbouring song sparrows. Animal Behaviour 51: 917-923.
Catchpole, C.K. & Slater, P.J.B. 1995. Bird song - biological themes and variations. Cambridge, Cambridge University Press.
Cody, M.L. & Brown, J.H. 1969. Song asynchrony in chapparal birds. Nature 222: 778-780.
Crowder, R.G. 1976. Principles of learning and Memory. Hillsdale, NJ, Lawrence Erlbaum.
Dobson, C.W. & Lemon, R.E. 1977. Markovian versus rhomboidal patterning in the song of Swainson's Thrush. Behaviour 62: 277-297.
Falls, J.B. 1985. Song matching in western Meadowlarks. Canadian Journal of Zoology 63: 2520-2524.
Falls, J.B. & d'Agincourt, L.G. 1982. Why do Meadowlarks switch song types? Canadian Journal of Zoology 60: 3400-3408.
Falls, J.B., Krebs, J.R. & McGregor, P.K. 1982. Song matching in the Great Tit (Parus major): the effect of similarity and familiarity. Animal Behaviour 30: 977-1009.
Falls, J.B., Horn, A.G. & Dickinson 1988. How Western Meadowlarks classify their songs: evidence from song matching. Animal Behaviour 36: 579-585.
Farabaugh, S.M. 1982. The ecological and social significance of duetting. In: Kroodsma, D.E. & Miller, E.H. (eds) Acoustic communication in birds 2: New York; Academic Press: 85-124.
Ficken, R.W., Ficken, M.S. & Hailman, J.P. 1974. Temporal pattern shift to avoid acoustic interference in singing birds. Science 183: 762-763.
Freyschmidt, J., Kopp, M.L. & Hultsch, H., 1984. Individuelle Entwicklung von gelernten Gesangsmustern bei Nachtigallen. Verhandlungen Deutsche Zoologische Ges. 77: 244.
Heupel, K. 1996. Ontogenese und tageszeitliche Variation der Zeitstruktur im Gesang der Singdrossel (Turdus Philomelos ). Diplom-Thesis Faculty of Biology, Free University of Berlin.
Hinde, R.A. 1958. Alternative motor patterns in Chaffinch song. Animal Behaviour 6: 211-218.
Horn, A.G. & Falls, J.B. 1986. Western meadowlarks switch son-types when matched by playback. Animal Behaviour 34: 927-929
Hultsch, H. 1980. Beziehungen zwischen Struktur, zeitlicher Variabilität und sozialem Einsatz im Gesang der Nachtigall (Luscinia megarhynchos). PhD Thesis, Free University of Berlin.
Hultsch, H. 1985. Formation of sub-repertoires: a strategy to support development and performance of large vocal repertoires. Verhandlungen Deutsche Zoologische Ges. 78: 229.
Hultsch, H. 1989. Ontogeny of song patterns and their performance mode in nightingales. In: Erber, J., Menzel, R., Pflüger, H.J. & Todt, D. (eds) Neural Mechanisms of Behaviour. Stuttgart; Thieme: p. 113.
Hultsch, H. 1990. Recombination of acquired songs as a correlate of package formation. In: Elsner, N. & Roth, G. (eds) Brain, perception, cognition. Stuttgart; Thieme: p. 433.
Hultsch, H. 1991a. Early experience can modify singing styles - evidence from experiments with Nightingales (Luscinia megarhynchos). Animal Behaviour 42: 883-889.
Hultsch, H. 1991b. Correlates of repertoire constriction in the song ontogeny of Nightingales (Luscinia megarhynchos). Verhandlungen Deutsche Zoologische Ges 84: 474.
Hultsch, H. 1991c. Song ontogeny in birds: closed or open developmental programs? In: Elsner, N. & Penzlin, E. (eds) Synapse, ransmission, modulation. Stuttgart; Thieme: p. 576.
Hultsch, H. 1992. Time window and unit capacity: dual constraints on the acquisition of serial information in songbirds. Journal Comparative Physiology A 170: 275-280.
Hultsch, H. 1993. Tracing the memory mechanisms in the song acquisition of birds. Netherland Journal of Zoology 43: 155-171.
Hultsch, H. & Todt, D. 1981. Repertoire sharing and song post distance in Nightingales. Behavioral Ecology and Sociobiology 8: 182-188.
Hultsch, H. & Todt, D. 1982. Temporal performance roles during vocal interactions in Nightingales. Behavioral Ecology Sociobiology 11: 253-260.
Hultsch, H. & Todt, D. 1986. Zeichenbildung durch mustergleiches Antworten. Zeitschrift für Semiotik 8: 233-244.
Hultsch, H. & Todt, D. 1989a. Song acquisition and acquisition constraints in the Nightingale (Luscinia megarhynchos). Naturwissenschaften 76: 83-86.
Hultsch, H. & Todt, D. 1989b. Memorization and reproduction of songs in Nightingales (Luscinia megarhynchos): Evidence for package formation. Journal Comparative Physiology A 165: 197-203.
Hultsch, H. & Todt, D. 1989c. Context memorization in the learning of birds. Naturwissenschaften 76: 584-586.
Hultsch, H. & Todt, D. 1992a. Acquisition of serial patterns in birds: temporal cues are mediators for song organisation. Bioacoustics 4: 61-62.
Hultsch, H. & Todt, D. 1992b. The serial order effect in the song acquisition of birds. Animal Behaviour 44: 590-592.
Hultsch, H. & Todt, D. 1996. Discontinuous and incremental processes in the song learning of birds: evidence for a primer effect. Journal Comparative Physiology A: 291-299.
Hultsch, H. Geberzahn, N. & Schleuss, F. 1998. Song invention in nightingales: Cues from song development. Ostrich 69: 254.
Hultsch, H., Lange, R. & Todt, D. 1984. Pattern-type labeled tutoring: a method for studying song-type memories in repertoire birds. Verhandlungen Deutsche Zoologische Ges. 77: p. 249.
Kopp, M.L. 1996. Ontogenetische Veränderungen in der Zeitstruktur des Gesangs der Nachtigall (Luscinia megarhynchos). PhD Thesis, Faculty of Biology, Free University of Berlin.
Kramer, H.G. & Lemon, R.E. 1983. Dynamics of territorial singing between neighbouring Song Sparrows (Melospiza melodia). Behaviour 85: 198-223.
Kramer, H.G. & Lemon, R.E. & Morris, M.J. 1985. Song switching and agonistic stimulation in the song sparrow (Melospiza melodia). Animal Behaviour 33, 135-149.
Krebs, J.R., Ashcroft, R. & Webber, M. 1978. Song repertoires and territory defence in the Great Tit. Nature 271: 539-542.
Kroodsma, D.E. 1979. Vocal dueling among male Marsh Wrens: Evidence for ritualized expressions of dominance / subordinance. Auk 98: 506-15.
Kroodsma, D.E. 1988. Song-types and their use: developmental flexibility of the male blue-winged warbler. Ethology 79: 235-47.
Kroodsma, D.E. 1996. Ecology of passerine song development. In: Kroodsma, D.E. & Miller, E.H. (eds). Ecology and evolution of communication. Ithaka, NY; Cornell University Press: 3-19
Kroodsma, D.E. & Miller, E.H. (eds) 1982. Acoustic communication in birds 2. New York; Academic Press.
Kroodsma, D.E. & Miller, E.H. (eds) 1996. Ecology and evolution of acoustic communication in birds. Ithaka, NY; Cornell University Press.
Lemon, R.E. 1968. Coordinated singing by Black-crested Titmice. Canadian Journal of Zoology 46: 1163-1167.
Marler, P. 1976. Sensory templates in species-specific behavior. In: Fentress, J.C. (ed) Simpler networks and behavior. Sunderland, MA, Sinauer Associates: 314-329.
Marler, P. 1987. Sensitive periods and the role of specific and general sensory stimulation in birdsong learning. In: Rauschecker, J.P. & Marler, P. (eds) Imprinting and cortical plasticity. New York; Wiley: 99-135.
Marler, P. 1991. Differences in behavioural development in closely related species: birdsong. In: Bateson, P.P.G. (ed) The development and integration of behaviour. Essays in the honour of Robert Hinde. Cambridge, Cambridge University Press: 41-70.
Marler, P. & Peters, S. 1982a. Structural changes in song ontogeny in the Swamp Sparrow (Melospiza georgiana). Auk 99: 446-58.
Marler, P. & Peters, S. 1982b. Developmental overproduction and selective attrition: New processes in the epigenesis of birdsong. Developmental Psychobiology 15: 369-78.
McGregor, P.K. 1991. The singer and the song: on the receiving end of bird song. Biological reviews 66: 57-81
McGregor, P.K. , Dabelsteen, T. Shephard, M & Pedersen, S.B. 1992. The signal value of matched singing in great tits: evidence from interactive playback experiments. Animal Behaviour 43: 987-998.
Morton, E.S. 1982. Grading, discreteness, redundancy and motivation-structural rules. In: Kroodsma, D.E. & Miller, E.H. (eds) Acoustic Communication in Birds 2. New York; Academic Press: 183-212.
Müller-Bröse, M. & Todt, D. 1991. Lokomotorische Aktivität von Nachtigallen (Luscinia megarhynchos) während auditorischer Stimulation mit Artgesang, präsentiert in ihrer lernsensiblen Altersphase. Verhandlungen Deutsche Zool. Ges. 84: 476- 477.
Naguib, M. 1997. Ranging of songs in Carolina wrens: effects of familiarity with the song-type on use of different cues. Behavioural Ecology and Sociobiology 40: 385-393.
Naguib, M. & Todt, D. 1997. Effects of dyadic vocal interactions on other conspecific receivers in Nightingales. Animal Behaviour 54: 1535-1543.
Nicolai, J. 1976. Beobachtungen an einigen paläarktischen Wintergästen in Ost-Nigeria. Die Vogelwarte 28: 274-278.
Nielsen, B.M.B. & Vehrencamp, S.L. 1995. Responses of Song Sparrows to song-type matching via interactive playback. Behavioural Ecology & Sociobiology 37: 109-117.
Shackleton, S.A. & Ratcliffe, L.. 1994. Matched counter-singing signals escalation of aggression in Black-capped Chickadees (Parus atricappillus) Ethology 97: 310-316.
Smith, D.G. & Reid, F.A. 1979. Roles of the song repertoire in Red-winged Blackbirds. Behavioural Ecology and Sociobiology 5: 279-290.
Slater, P.J.B. 1978. A simple model for competition between behaviour patterns. Behaviour 67: 236-257.
Slater, P.J.B. 1983. Sequences of songs in Chaffinches. Animal Behaviour 31: 272-281.
Slater, P.J.B. 1989. Bird song learning: causes and consequences. Ethology Ecology & Evolution 1: 19-46.
Thimm, F.A., Clausen, A., Todt, D. & Wolffgramm, J. 1974. Zeitabhängigkeit und Verhaltensmusterfolgen. Journal Comparative Physiology 93: 55-84.
Todt, D. 1968. Zur Steuerung unregelmässiger Verhaltensabläufe. In: Mittelstaedt, H. (ed) Kybernetik. Oldenbourg, München: 465-485.
Todt, D. 1970a. Gesang und gesangliche Korrespondenz der Amsel. Naturwissenschaften 57: 61-66.
Todt, D. 1970b. Gesangliche Reaktionen der Amsel auf ihren experimentell reproduzierten Eigengesang. Zeitschrift Vergleichende Physiologie 66: 294-317.
Todt, D. 1970c. Zur Ordnung im Gesang der Nachtigall (Luscinia megarhynchos). Verhandlungen Deutsche Zool. Ges. 64: 249-252.
Todt, D. 1971a. Äquivalente und konvalente gesangliche Reaktionen einer extrem regelmässig singenden Nachtigall (Luscinia megarhynchos B.). Zeitschrift Vergleichende Physiologie 71: 262-285.
Todt, D. 1971b. Abstraktionsleistung einer gesanglich korrespondierenden abessinischen Erddrossel (Geokichla piagiae). Zeitschrift für Tierpsychologie 28: 59-61.
Todt, D. 1974. Social learning of vocal patterns and modes of their application in Grey Parrots (Psittacus erithacus). Zeitschrift für Tierpsychologie 39: 178-188.
Todt, D. 1975. Short term inhibition of vocal outputs occurring in the singing behaviour of blackbirds (Turdus merula). Journal Comparative Physiology 98: 289-306.
Todt, D. 1977. Zur infradianen Rhythmik im Verhalten von Vertebraten-Ergebnisse aus Analysen des variablen Verhaltens von Singvögeln. Nova Acta Leopold 225: 607-619.
Todt, D. 1981. On functions of vocal matching: effect of counter-replies on song-post choice and singing. Zeitschrift für Tierpsychologie 57: 73-93.
Todt, D. & Böhner, J. 1994. Former experience can modify social selectivity during song learning in the nightingale (Luscinia megarhynchos). Ethology 97: 169-176.
Todt, D. & Fiebelkorn, A. 1979. Display, timing and function of wing movements accompanying duets of Cichladusa guttata. Behaviour 25: 42-58.
Todt, D. & Hultsch, H. 1980. Functional aspects of sequences and hierarchy in bird song. Acta XVII. Congressus Intern. Ornithologici, Berlin: 663-670.
Todt, D. & Hultsch, H. 1994. Biologische Grundlagen des Dialogs. In: Wessel, K.F & Naumann, F. (eds). Kommunikation und Humanontogenese. Bielefeld, Kleine Verlag: 53-76.
Todt, D. & Hultsch, H. 1996. Acquisition and performance of repertoires: ways of coping with diversity and versatility. In: Kroodsma, D.E. & Miller, E.H. (eds). Ecology and evolution of communication. Ithaka, NY; Cornell University Press: 79-6.
Todt, D. & Wolffgramm, J. 1975. Überprüfung von Steuerungssystemen zur Strophenwahl der Amsel durch digitale Stimulierung. Biol. Cybern. 17: 109-127.
Todt, D., Hultsch, H. & Duvall II, F.P. 1981. Behavioural Significance and Social Function of Vocal and Non-Vocal Displays in the Monogamous Duet-Singer (Cossypha heuglini H). In: Strübing, H. (ed) Zoologische Beiträge. Berlin; Duncker & Humblot: 422-448.
Todt, D., Hultsch, H. & Heike, D. 1979. Conditions affecting song acquisition in Nightingales (Luscinia megarhynchos). Zeitschrift für Tierpsychologie 51: 23-25.
Wasserman, F.E. 1977. Intraspecific acoustical interference in the White-throated Sparrow (Zonotrichia albicollis). Animal Behaviour 25: 949-952.
Wistel-Wozniak, A. & Hultsch, H. 1992. Song performance in Nightingales which had been raised without exposure to acoustic learning programmes. Verhandlungen Deutsche Zoologische Ges. 85: 246.
Wistel-Wozniak, A. & Hultsch, H. 1993. Konstante und altersabhängig veränderte Gesangsmerkmale bei handaufgezogenen Nachtigallen. Verhandlungen Deutsche Zoologische Ges. 86: 281.
Whitney, C.L. 1981. Patterns of singing in the Varied Thrush. II. A model of control. Zeitschrift für Tierpsychologie 57: 141-162.
Wolffgramm, J. & Todt, D. 1982. Pattern and time specificity in vocal responses of Blackbirds (Turdus merula). Behaviour 81/2-4: 264-286.
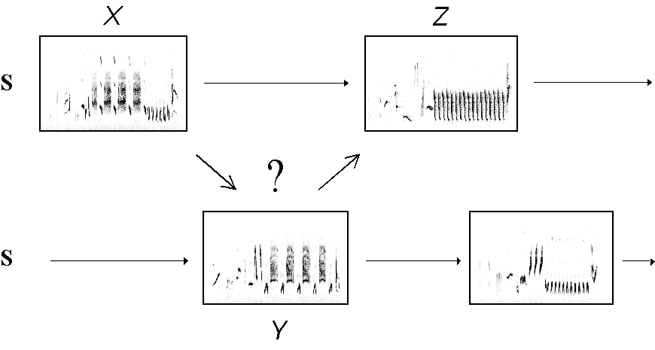
Fig. 1. Spectrograms of three song-types (X, Y, Z) produced by two neighbouring Nightingales. We investigate time and patterns specific relationships between exchanged songs. (after Todt & Hultsch 1994)
Fig. 2. Frequency histograms of intersong interval durations assessed in a Nightingale, who was exposed to interactive playback experiments during his nocturnal singing. The top histogram gives interval durations in his spontaneous solo-singing, the lower ones those assessed during the stimulation. Inserted horizontal bars indicate the onset and duration of the stimulus songs. The onset of stimuli refers to ‘Zero’, which is the time when the bird had terminated a given song. (after Hultsch 1980)
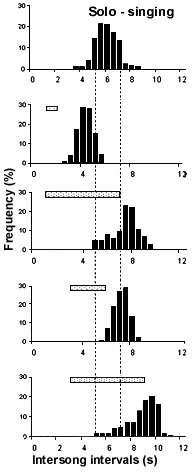
Fig. 3. Intersong intervals (median, 1st & 3rd quartile) of the tested male plotted against the number of songs he had produced after the end of a playback trial. (after Hultsch 1980)
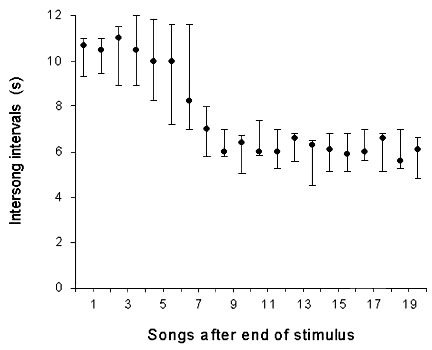
Fig. 4. Acquisition success of master songs plotted against the number of times birds were allowed to hear these songs during the tutoring.
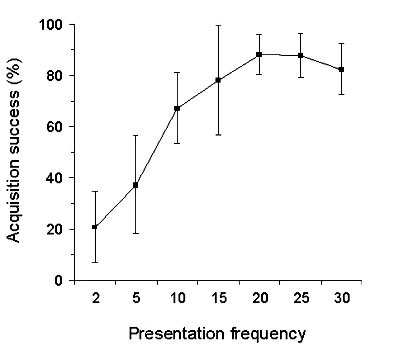
Fig. 5. Illustration of a learning experiment with two social tutors (one familiar, one strange). In an alternating presentation, either tutor played an ‘individual’ set of master songs.
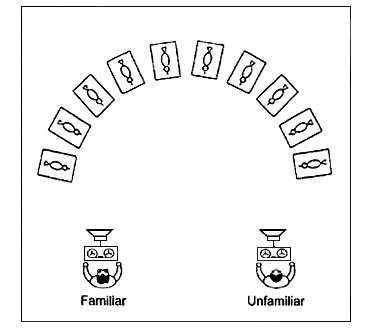
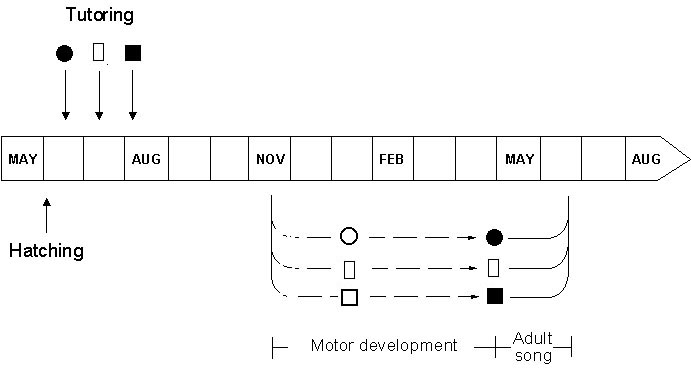
Fig. 6. Illustration of the time course of vocal development in a handraised Nightingale.