S03.2: Social interactions and song learning: Are behavioural contingencies important?
Carel ten Cate & Bart B. Houx
Behavioural Biology, Institute of Evolutionary and Ecological Sciences, Leiden University, PO Box 9516, 2300 RA Leiden, The Netherlands, fax 31 71 5274900, e-mail tencate@rulsfb.leidenuniv.nl
ten Cate, C. & Houx, B.B. 1999. Social interactions and song learning: Are behavioural contingencies important? In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban:
156-164. Johannesburg: BirdLife South Africa.Several songbird species are known to copy their song more easily from a live tutor than from playback. Social interactions between tutor and pupil are assumed to be responsible for this finding. These interactions may provide a context which stimulates the learning process. We present the results of several experiments on song learning in Zebra finches Taeniopygia guttata, aimed at testing the hypothesis that contingencies in the interaction between tutor and pupil are the key factor underlying the effect of interactions. The results show that there are no strong contingencies between singing of the tutor and pupil behaviour during normal rearing. Exposing isolated Zebra finch pupils to playback which is either contingent upon behaviour of the pupil (key pecking) or other events (presentation of a stuffed Zebra finch) does lead to copying of the song, but no more so than in controls lacking any contingencies. The results cast doubt on the contingency hypothesis. However, they do show that the Zebra finch's ability to copy from playback is better than previous studies indicated.
INTRODUCTION
Songbirds generally learn their songs by copying from a 'tutor'. Some species, like the White-crowned Sparrow Zonotrichia leucophrys (Marler 1970) are able to imitate songs from playback. However, over the last decade it has become increasingly clear that not just exposure to song is important for copying, but also the way in which the juvenile is exposed to this song. In particular, exposure to, and interaction with, a live tutor improves song copying. For instance, in the white-crowned sparrow exposure to a live tutor may facilitate song learning and may even induce copying of elements of allospecific songs (Baptista & Petrinovich 1986). In other species, too, song copying is facilitated by or requires exposure to a live tutor. A species for which such 'social tutoring' has been considered essential for any learning to occur is the Zebra finch. Live tutoring has been shown to induce copying of tutor songs, not only of conspecific, but also of allospecific songs (Immelmann 1969). In contrast, tutoring by playback (ten Cate 1991) or through an audiolink (Eales 1989) led to no or very little copying. The Zebra finch has become a model species for investigating which aspects of interactions with live tutors may be important and, also, what mechanisms might underlie the effects. Observations of Zebra finch tutors and pupils during the sensitive phase for song learning showed that the degree to which a tutor song was copied was related to factors like the amount of interaction (Williams 1990), proximity to the tutor (Mann & Slater 1995) or tutor aggression (Clayton 1987, Jones & Slater 1996). However, not all studies showed the same effects and the results are not clear cut. Also, even if a particular behaviour pattern 'X' does show a strong correlation with copying, the question of the mechanisms underlying such an effect remains: what causes song copying from a tutor performing X (ten Cate 1994; Nelson 1997)? It is this question we address in the experiments reported in this paper.
One suggestion for how social interactions exert their effect is the 'contingency hypothesis'. This hypothesis was put forward by ten Cate (1991, 1994), based on experiments on sexual imprinting, which demonstrated that the amount of learning about an imprinting model was correlated less with the amount of visual exposure to this model than with the number and nature of behavioural interactions (ten Cate 1989). Here too, various behaviour patterns seemed to contribute to the effect. Ten Cate suggested that the result might have been obtained because the interactions may have involved contingencies between the behaviour of the model and being exposed to its appearance (ten Cate 1989, 1994). Such contingencies might have been crucial for learning to take place. This contingency hypothesis has been tested for filial imprinting, with ambiguous results (ten Cate 1986; Bolhuis & Johnson 1988). Ten Cate suggested that the hypothesis might also apply to song learning: When singing of the tutor is associated with meaningful behaviour of the tutor or is a response to behaviour of the pupil, these patterns provide contingencies which in turn might enhance song learning. The hypothesis led to an experiment, in which exposure to song was made contingent upon perch hopping in Zebra finches (ten Cate 1991). The results showed that, in spite of a significant increase in hopping on the reinforced perch, young males did not copy the playback song. However, in a subsequent experiment, Adret (1993), using key pecking as the operant, obtained evidence that the males controlling the exposure to song copied from the playback and more so than their yoked controls. However, the experimental bird and the yoked control were in the same room and could interact vocally. Therefore, ascribing the full effect to the presence of a contingency seems premature and the results thus provide no strong test of the contingency hypothesis.
In this paper we describe three experiments aimed at a more rigorous analysis of the effect of contingency in song learning in Zebra finch males. We can envisage three (not mutually exclusive) types of contingencies between song of the tutor and other behaviour which may be important in promoting song learning (see also Houx & ten Cate 1998): (1) Behaviour of the tutor may predict subsequent singing; (2) singing of the tutor may predict meaningful tutor behaviour (e.g. aggression); and (3) behaviour of the juvenile may stimulate singing by the tutor. Of these contingencies, the first two are classical or stimulus-stimulus associations, the third one is an operant association.
In our first experiment we observed the behavioural interactions between fathers and their male offspring. In this way we examined whether contingencies of the above mentioned types do occur. The second experiment concentrated on the possible effect of operant contingencies by examining whether song contingent upon key pecking leads to enhanced song learning. In this experiment the possibilities for interaction between experimentals and yoked controls, as may have occurred in the earlier experiments of Adret (1993), were excluded. The third experiment tested the possibility that classical (stimulus-stimulus) contingencies influence the learning process, by comparing the song learning in pupils exposed to various associations between song playback and visual stimulation with a stuffed model. The potential for such stimulus combinations to enhance vocal learning was demonstrated by van Kampen & Bolhuis (1991). They showed that young chicks developed a stronger preference for an auditory stimulus when this was combined with exposure to a visual stimulus compared to when the auditory stimulus was presented on its own. Below, the experiments and results will be described one by one.
CONTINGENCIES DURING NORMAL REARING
The details and full data of this experiment are published elsewhere (Houx & ten Cate 1998), hence we only provide the most important aspects of methods and results.
Methods
Six pairs of Zebra finches with their broods were observed. Brood-size was reduced to three at the start of the observations, around day 30 after hatching. In each pair but one, the adult male and two juvenile males served as focal animals up to around day 66. In one pair there was only one juvenile male; this male and his father served as focal animals in this pair. Each focal animal was observed during five sessions per week, for thirty minutes per session. The various behaviour patterns included all regularly observable social behaviour patterns of the focal animal and to whom they were directed (e.g. approaching/leaving, allopreening, various aggressive and sexual behaviours, singing).
In order to investigate whether a particular behaviour pattern was contingent with undirected or directed singing of the tutor, we analysed whether the different behaviour patterns of the tutor occurred more (or less) than expected by chance, before or after tutor singing. We also examined whether singing of the tutor was preceded by any particular pupil behaviour at more than chance level. To this end we calculated observed and expected frequencies of behaviour patterns of the pupil for 10 sec before and 15 sec after the start of tutor singing.
Results
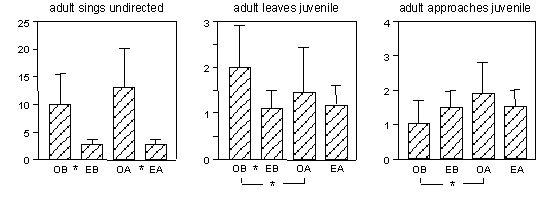
The mean frequencies of undirected and directed singing of the fathers were 6.2 and 0.9 times per 30 min period. For both types of singing very few indications were present that a particular behaviour reliably either preceded (i.e. signalled) or followed (i.e. was signalled by) singing. Fig. 1 shows all behaviour patterns of the tutors for which there was either a significant difference between the observed and expected frequency of undirected song or for which the values before and after singing were significantly different. Directed singing was preceded more by undirected singing than expected, but apart from that no relation between directed song and tutor behaviour was present. Juvenile males showed less singing than expected before undirected or directed tutor song, but no other behaviour deviated from the expected value. All males showed considerable matching between their own song and that of their father (79.6%, S.D. 16.4), showing that copying had occurred.
Discussion
No clear contingencies were present between singing of the tutor and his own behaviour or that of the pupil preceding tutor song. This was unexpected in view of the studies mentioned above, which showed correlations between the overall level of some behavioural interactions and the degree of song copying (Clayton 1987; Williams 1990; Mann & Slater 1995; Jones & Slater 1996) and also in view of Adret’s study (Adret 1993) which indicated an effect of contingency on song copying. Our results may indicate that such correlations most likely do not result from immediate contingencies between performing the behaviour and singing. Rather, interactions might provide a general change in motivational state which might make copying more likely to occur. Contrary to our expectations this may be taken to suggest a limited role for direct contingencies for song copying.
OPERANT CONTINGENCIES AND SONG COPYING
This experiment examined whether song copying was stimulated when exposure to playback of song contingent upon performing an operant (key pecking).
Methods
Two groups of eight males each were used (details concerning this experiment are given in Houx & ten Cate (1999a)). All males had their father removed around the age of 8 days. They could, however, hear other adult males. Between 30 and 65 days of age each young male was isolated from others in a Skinner box in a sound attenuated room. The box contained five perches, two of which were in front of an illuminated pecking key. One group (experimental males) consisted of males for which a peck on one of the keys led to playback of five sec of adult male song. Each of the experimental males was linked to a yoked control in an adjacent room. This male had no control over the exposure, but received the same song at the same moment as the experimental male. Each set of one experimental and its yoked control male received a different song. The songs originated from males of the same breeding colony. After 65 days the males were removed from the Skinner boxes and kept isolated from other finches. The songs of all males were recorded when adult (average age of recording 131 days). The song of each male was analysed and compared with that of his tutor as well as with one randomly selected unfamiliar song (which was a tutor song for another pupil). The difference between these two was taken as a measure for how much the pupils learned from the playback. Song copying of experimental males was compared with that of their yoked controls. The degree of song matching was scored by three independent observers, who were unaware of the experimental history of the different songs.
Results
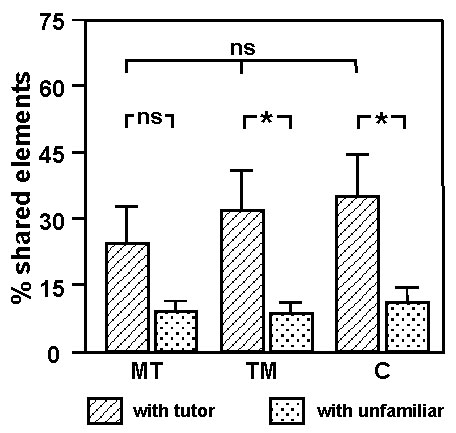
All young males pecked more to the reinforced key than to the non-reinforced one, indicating that hearing song was reinforcing. Over the last eight days in the Skinner box the level of pecking was on average 280 pecks per day (S.D. 123). Fig. 2 presents the results of both experimental and control males. Both showed a significantly greater similarity of their song to the tutor song than to the unfamiliar song (Wilcoxon test, P<0.05). Between groups there were no significant differences in similarity to either tutor song or unfamiliar song.
Discussion
This experiment confirms earlier findings that song can act as a reinforcer in young Zebra finch males (ten Cate 1991; Adret 1993). Our study also confirms that Zebra finches may copy their song from playback (Adret 1993). However, in contrast to Adret, we found no evidence that a contingency between key pecking and playback improved song copying. A reason for the discrepancy between our results and those of Adret may be that we prevented vocal interactions between the experimental and the control male. However, it must also be noted that Adret's sample of birds was small (N=3) and need not have been representative. Whatever the cause of the differences, our data suggest that we have to reject the possibility that an operant contingency between pupil behaviour and song playback is responsible, or necessary, for copying. The reason that earlier studies did not obtain evidence that Zebra finch males can learn from playback (Eales 1989; ten Cate 1991) while we do, remains unclear. However, we have some indications (Houx, unpublished) that the sound quality of the playback may affect whether or not a song is copied.
CLASSICAL CONTINGENCIES AND SONG COPYING
This experiment examined whether stimulus-stimulus contingencies facilitate song copying. Song was used either as a stimulus which signalled the appearance of a stuffed male (see Houx & ten Cate, 1999b) Zebra finch, or as a stimulus whose playback was signalled by the appearance of a stuffed male.
Methods
Three groups of eight males each were used in this experiment. All males were treated as those in the previous experiment, apart from the tutoring period in the Skinner box. When in this box, keys were switched off. Instead, the computer device presented the birds with playback of tutor song using a quasi-random schedule giving 20 presentations per hour. For one group (TM) playback of tutor (T) song was followed by 6 sec exposure to a stuffed male (=model; M). In the second group (MT) the sequence of exposure to song and model was reversed. A third group served as control; here song was played, but no model was presented. The level of exposure to song was, on average, higher for the males in the present experiment than it was for the groups in the operant experiment. We used the same songs as in the previous experiments, which means that all males within a group were exposed to a different song and that each song was presented to one TM male, one MT male and one control male. As in the experiments presented above, pupil songs were recorded at approximately 130 days of age and compared with the tutor song and an unfamiliar song.
Results
Fig. 3 shows that the songs of all three groups had a greater similarity to the tutor song than to the unfamiliar one, although this difference was not significant in the MT group. There were no significant differences between the three groups. As the same songs were used in this experiment and in the previous one, we compared for each song whether the mean value of matching obtained by the experimental bird and his yoked control in the operant experiment differed from the mean of the three birds (TM, MT, C) exposed to that same song in the present experiment. The value of the males from the operant experiment was significantly higher (Wilcoxon test, P<0.05), indicating that these males copied more than the males from the classical contingency experiment. However, the difference between the control groups of both experiments was not significant.
Discussion
Like the previous experiment, this one shows that Zebra finch males can copy song from playback, albeit at a modest level in this case. In addition, as in the previous experiment, the findings show no evidence that exposure to song contingent with exposure to a model (TM and MT groups) gives rise to a higher degree of copying than exposure to song which is not associated with presentation of a visual stimulus. So, here again no effect of contingency was obtained. An explanation for the higher amount of copying in the operant experiment compared with the classical experiment might be the pattern of exposure to song. That is, in the operant experiment song was presented when the experimental bird pecked, and this led to a pattern in which exposure to song was higher in the mornings and also higher toward the end of the period in the Skinner boxes. In the present experiment song exposure was spread evenly between and within days. It might be that in the latter schedule exposure to song coincided less with the periods during which Zebra finch males are most motivated to listen to and learn about song.
GENERAL DISCUSSION
Our experiments do not provide evidence to support the hypothesis that behavioural contingencies in relation to exposure to song would enhance song copying. What our experiments show is that Zebra finches can copy song from playback, even if this playback is not accompanied by any other event. So, in contrast to earlier findings (Eales 1989; ten Cate 1991), our data show that social interaction is not a crucial feature of the song learning process in Zebra finches. At the same time, however, the data indicate that copying in the classical condition is lower than in the operant one, and in the latter condition lower than with live tutors. This last finding may indicate that social interaction does enhance copying, for instance through a motivational effect on the young bird. However, the difference between the two contingency experiments indicates that seemingly trivial details such as timing of exposure to song might matter. If so, there might be other aspects than social interaction per se which make live tutors the most effective ones (e.g. song by live tutors may be more variable than playback song in subsequent renditions, preventing habituation). Our study highlights that such possibilities should not be overlooked or abandoned too easily for more complex or cognitive explanations. For the moment we can only conclude that whatever aspect might make live tutors most effective, contingencies between song output of the tutor and his or the pupils behaviour are not a factor of importance.
ACKNOWLEDGEMENTS
We thank Enja Feuth-de Bruijn for her help in the various experiments, Juliette Bergfeld for looking after the birds, Rinus Heijmans, Ab Gluvers, Rob van der Linden and their collegues for building the Skinner boxes and Johan Bolhuis and Irene Pepperberg for their comments on the manuscript.
REFERENCES
Adret, P. 1993. Operant conditioning, song learning and imprinting to taped song in the Zebra finch. Animal Behaviour 46: 149-159.
Baptista, L.F. & Petrinovich, L. 1986. Song development in the white crowned sparrow: Social factors and sex differences. Animal Behaviour 32: 172-181.
Bolhuis, J.J. & Johnson, M.H. 1988. Effects of response-contingency and stimulus presentation schedule on imprinting in the chick (Gallus gallus domesticus). Journal of Comparative Psychology 102: 61-65.
Clayton, N.S. 1987. Song tutor choice in Zebra finches. Animal Behaviour 35: 714-721.
Eales, L.A. 1989. The influences of visual and vocal interaction on song learning in Zebra finches. Animal Behaviour 37: 507-508.
Houx, B.B. & ten Cate, C. 1999a. Song learning from playback in Zebra finches: is there an effect of operant contingency? Animal Behaviour 57: in press.
Houx, B.B. & ten Cate, C. 1999b. Do stimulus-stimulus contingencies affect song learning in Zebra finches (Taeniopygia guttata)? Journal of Comparative Psychology: in press.
Houx, B.B. & ten Cate, C. 1998. Do contingencies with tutor behaviour influence song learning in Zebra finches? Behaviour: 135: 599-614.
Immelman, K. 1969. Song development in the Zebra finch and other Estrildid finches. In: Hinde, R.A. (ed.). Bird vocalizations. Cambridge University Press; Cambridge: 61-74.
Jones, A.E. & Slater, P.J.B. 1996. The role of aggression in song tutor choice in Zebra finches: cause or effect? Behaviour 133: 103-115.
Mann, N.I. & Slater, P.J.B. 1995. Song tutor choice by Zebra finches in avaries. Animal Behaviour 49: 811-820.
Marler, P. 1970. A comparative approach to vocal learning: song development in the white crowned sparrows. Journal of Comparative and Physiological Psychology, 53:297-301.
Nelson, D.A. 1997. Social interaction and sensitive ophases for song learning. A critical review. In: Snowdon, C.T. & Hausberger, M.E. (eds.) Social influences on vocal development. Cambridge University Press; Cambridge: 7-22.
ten Cate, C. 1986. Does behaviour contingent stimulus movement enhance filial imprinting in Japanese quail? Developmental Psychobiology 19: 263-285.
ten Cate, C. 1989. Behavioral development: towards understanding processes. In: Bateson, P.P.G. and Klopfer, P.H. (eds.) Perspectives in Ethology, vol. 8; New York; Plenum Press: 243-269.
ten Cate, C. 1991. Behaviour-contingent exposure to song and Zebra finch song learning. Animal Behaviour 42: 857-859.
ten Cate, C. 1994. Perceptual mechanisms in imprinting and song learning. In: Hogan, J.A. & Bolhuis, J.J.(eds.) Causal mechanisms of behavioural development. Cambridge University Press; Cambridge: 116-146.
van Kampen, H.S. & Bolhuis, J.J. 1991. Auditory learning and filial imprinting in the chick. Behaviour 117: 303-319.
Williams, H. 1990. Models for song learning in the Zebra finch: fathers or others? Animal Behaviour 39: 745-757.
Fig. 1. Percentage of times (with S.D.) that the onset of undirected song of the adult males is preceded or followed by specific behaviour patterns of those males. OB = observed during the ten seconds before song; EB = expected value for this period; OA = observed during the 15 seconds after song onset; EA = expected value for this period. * indicate significant differences (P<0.05).
Fig. 2. Percentage of the elements of experimental and yoked control birds which they shared with their (playback) tutor song and with an unfamiliar song. * = significant difference (P<0.05) with a Wilcoxon test.
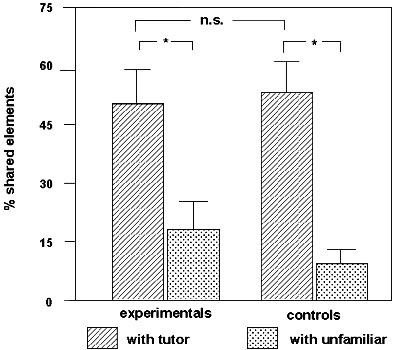
Fig. 3. Percentage of the elements of the MT, TM and C birds which they shared with the tutor song and with an unfamiliar song. * = significant difference (P<0.05) with a Wilcoxon test