S03.1: Cognitive Processes in Avian Vocal Development
Luis F. Baptista1 & Sandra L.L. Gaunt2*
1California Academy of Sciences, Golden Gate Park, San Francisco, CA, USA, e-mail lbaptista@casmail.calacademy.org; 2Borror Laboratory of Bioacoustics, Ohio State University, Columbus, Ohio, USA, fax 614 292 7774, e-mail gaunt.2@osu.edu; *Author for correspondenceBaptista, L.E., Gaunt, S.L.L. 1999. Cognitive processes in avian vocal development. In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 138-155. Johannesburg: BirdLife South Africa.
Although cognition is difficult to demonstrate in any animal species, observations and experiments may elucidate cognitive processes associated with avian vocal learning. These processes include memory, selective attention, categorisation, information processing, and decision-making. Each of these processes is involved to some extent with the two phenomena that have been proposed to constitute avian vocal learning: learning (storage) for discrimination and learning for performance. Playback studies have however demonstrated that vocal learning involves a continuum between these phenomena, with storage and discrimination at one end, storage, discrimination, and performance at the other, and storage, discrimination, and occasional performance in between. We examine how cognitive processes are involved in the behaviour of several species with respect to these learning phenomena.
INTRODUCTION
General cognition, the selection, retention and later use of retained stimuli from memory, can be difficult to demonstrate. ‘Domain specific’ or specialised cognitive processes, such as vocal learning, have, in contrast, proven amenable to study and have contributed much to our understanding of cognition (Kamil 1994, Moscovitch 1995). Avian vocal learning studies have been especially insightful in elucidating cognitive processes; research has been both observational and experimental, involving a wide spectrum of behavioural phenomena from the timing of and mechanisms for acquiring vocal memories to the retrieval of stored vocal information used in social interactions.
Vocal learning involves cognitive processes of selective attention, memory retention, information processing and decision making and includes two, not mutually exclusive, uses of memory: memory storage for discrimination and storage for performance (McGregor & Avery 1986). In storage for discrimination, avian subjects assimilate auditory stimuli from the environment that emanate, usually, from members of their own species. Even if the subjects never produce the stored sound stimuli (i.e. their produced vocalisations are not learned, they are time or context restricted, or they are young birds in the early stage of vocal acquisition), they are nonetheless capable of discriminating between utterances produced by different individuals as a consequence of acquired memory. Storage for performance is well known in three groups of birds, namely the parrots (Psittacines), hummingbirds (Trochilidae) and true song-birds (Oscines) within the perching bird clade (Passeriformes). In storage for performance, naive juveniles, after assimilating auditory stimuli, vocalise accurate copies of some or all of those stimuli. The process from acquisition to production has been extensively studied experimentally in various avian species. The usual protocol involves exposing naive juveniles to acoustic models (tutors), as either a playback of vocalisation on tape or exposure to singing adult males, and then following the sound produced to maturity when accurate vocal copies of the model are performed ( Marler 1987, Baptista 1996, Baptista & Gaunt 1997a).
Indeed, such playback studies indicate that there is a continuum between species in how stored sound memories are used, with discrimination at one end and discrimination and performance at the other. In between are avian species that store, discriminate and occasionally vocalise (perform) stored vocalisations. This continuum can best be appreciated when we examine categorisation, retrieval and discrimination processes in both Oscines with learned performance and non-Oscines with ‘innate’ performance. In this context, we review cognitive processes associated with recognition of individuals, populations and allospecies. Individual recognition and social interactions are considered for numerous dyads including those between young and adults, adults and young, and between adults of the same and different sexes with particular emphasis on mate recognition. In the discussion we present the adaptive significance of these different aspects of vocal acquisition, storage and use.
STORAGE FOR DISCRIMINATION ONLY
Individual recognition
The ability to recognise and respond appropriately to specific members of your own species, group and/or family can be critical to survival. Nowhere is this more evident than in colonial or semi-colonial species, especially those that nest in mixed species aggregations. Under such conditions is a premium is placed upon juveniles to learn their parents' voices and upon mates to recognise one another.
Juvenile-parent recognition
Among non-Oscines, juvenile birds recognise parents in many species including alcids, gulls, terns, penguins (Falls 1982) and a dove species (Hitchcock et al. 1989). In the colonially breeding Common Murre Uria aalga, learning of parental contact calls by chicks is prenatal. The characteristic calls of parents are vocalised to the chicks in the egg at pipping, at which time the chicks commit the unique parental call to memory. Hatched chicks exposed to two speakers, one playing calls heard while in the egg and another broadcasting a strange call, approached the speaker playing the call heard prenatally (Tschanz 1968). Prenatal learning of parental calls has also been demonstrated in Wood Ducks (Aix sponsa, Miller & Gottlieb 1976) and Mallards Anas platyrhynchos (Johnston 1988). Mothers’ vocalise to newly hatched ducklings and, having already learned to recognise her call, the brood leaves the nest synchronously. The male of the altricial Mourning Dove Zenaida macroura vocalises to his nestlings during feeding bouts (Luther 1979). There is individual variation in songs of Morning Doves (Fig. 1), and playback studies indicate that fledglings may recognise their father's song (Hitchcock et al. 1989). Broods in Mourning Doves often overlap so that females are already incubating a new set of eggs by the time her chicks have fledged (Blockstein & Westmoreland 1993). The male then does the feeding so that it behoves his young to recognise his voice in this semi-colonial breeding species.
Young Oscines also learn to recognise parents, and adults can recognise their brood's vocalisations. Playback experiments demonstrated that fledglings of the semi-colonial Zebra Finch Taeniopygia guttata recognise the distance calls of parents and conversely parents recognise distance calls of fledglings (McIntosh 1983, cited in Zann 1996). Parents responded to calls of broods 20 days of age, the response waning when the young reach 32 to 35 days of age, ceasing completely beyond that age. Field studies revealed that five to seven days after fledging young would call to adults and could recognise parental calls up to 100 m away (Immelmann 1962). Playback experiments also indicated that female Zebra Finches could recognise their fathers' calls after many months of separation (Zann 1996). Memorisation of father's song for recognition commences when fledglings are 25 to 35 days old (Clayton 1988). Playback studies indicate that parents of colonially breeding Bank Swallows Riparia riparia and Cliff Swallows Hirundo pyrrhonota recognise calls of their fledged broods (Beecher et al. 1981, Stoddard & Beecher 1983).
Individual recognition is also important in non-colonial species, notably during brood division. Broods of fledgling Song Sparrows Melospiza melodia are divided between the parents, half the brood following the father and the other half following the mother. Spectrograms reveal that contact calls of young are individually distinct, enabling each parent to recognise the fledglings under their care (Smith & Merkt 1980). Fledgling Piñon Jays Gymnorhinus cyanocephala, another non-colonial species, join creches after fledging at 21 days of age. Nest exchange experiments indicate that adults and young learn to recognise each other's calls after day 14 when they develop individually distinct begging calls (Balda & Balda 1978, McArthur 1982).
Mate recognition
Experienced mated pairs are often more reproductively successful than newly mated pairs (review in Farabaugh & Dooling 1996) and thus recognition of a mate can be highly adaptive. Vocal signals may simply aid in the identification of an individual amongst many, as in the congested colonial breeding environment. However, recognition may also facilitate those species wherein mates co-operate in territorial defense and predator detection, or when males assist in the reproductive effort by incubating and/or feeding.
Two examples from the many possible within colonial breeding non-Oscines are presented. Emperor penguins Aptenodytes forsteri are not only colonial breeders, but have a peculiar habit of moving their single egg or chick from locality to locality. Mates cannot predict, on the basis of nest site alone, where its mate might reside, and it is thus not surprising that recognition of a mate's call has been documented for this species (Jouventin et al. 1979). The calls of colonial Petrels (Hydrobatidae) develop independently of imitation learning, yet sexual dimorphism and individual variation exists in call structures used in mate recognition (Bretagnolle 1996). This behaviour is especially critical in these nocturnal, burrowing species where auditory cues from the mate assist in burrow location.
Amongst colonial Oscines, Zebra Finch males have contact calls that are individually distinct. Playback studies indicate that mated pairs may recognise each other's calls (McIntosh 1983, cited in Zann 1996). Playback studies have also shown that Zebra Finch females may distinguish between their mate's song and the song of other males (Miller 1979).
Solitary breeding birds also recognise mates by vocal cues. Mate recognition by call has been documented in California Quail Lophortyx californica, Northern Bobwhite Colinus virginianus and Red-headed Woodpeckers Melanerpes erythricephalus (Williams 1969, Stokes 1967, review in Falls 1982). For example, the source of individual variation in the California Quail separation call is in the frequency of the first syllable (Williams 1969).
Species and population recognition
Birds, not unlike primates, preferentially attend to the vocalisations of their own species with vocal learners assimilate own species vocalisations more readily than other sounds in their environment. Examples of the capacity to recognise own species are, however, also available from non-Oscines.
Many colonial seabirds recognise the calls of their own species emanating from existing colonies and choose to breed at such sites. Conservation biologists have used this knowledge to establish or re-establish colonies on suitable new or historic breeding grounds that were abandoned, due to climactic events or human activity. The protocol involves coupling the use of decoys with the broadcast of the colony sounds (Kress & Nettleship 1988, Podolsky 1990, Podolsky & Kress 1992, Kress 1983).
Bobwhite Quail Colinus virginianus exhibit geographical variation in separation calls (Goldstein 1978). Coveys may also be distinguished from each other on the basis of separation call and cross-fostering experiments indicate that this variation is inherited (Baker & Bailey 1987a). However, chicks preferentially approached playback of calls of foster parents or covey mates with whom they were raised, indicating that youngsters learned to recognise individuals or groups by calls, although they themselves never produced them (Baker & Bailey 1987b).
Most female songbirds studied to date do not sing, but there is mounting evidence that they discriminate species, subspecies or population specific song. Playback studies with female Zebra Finches implanted with silastic tubes filled with estradiol revealed that they are capable of distinguishing between mainland versus insular songs. They performed more copulation solicitation displays to familiar songs than to alien songs (Clayton 1990 a,b). Female White-crowned Sparrows implanted with estradiol have been shown to distinguish between various song dialects (Baker et al. 1982, Chilton & Lein 1996).
Similarly, female Brown-headed Cowbirds Molothrus ater discriminate song at species and subspecies levels with preference being for songs of local, dominant males, and preference is exhibited as a social, copulation solicitation, displays to male with preferred song (King & West 1990). More intriguing in the Brown-headed Cowbird is the fact that females can channel development of male song learning behaviour; females give subtle wing signals to consubspecific males that are singing preferred song types, and males are attentive to these signals during the song learning process, and respond by modifying their songs to that type (King & West 1988). Significantly, the same brain areas used by males to acquire song (nucleus 1MAN) are found to be responsible for female Brown-headed Cowbird song acquisition, and females that are more selective in their male song preferences have larger 1MAN nuclie (Hamilton et al. 1997). Though it is clear that females show preferences for specific stimuli, how they store and categorise perceived song without the feedback from vocalising has yet to be discerned.
Allospecies recognition
Bird vocalisations have been so thoroughly documented to function in intraspecific territorial and mate advertisement, that their function interspecifically has been until recently largely overlooked. However, in a study of captive Darwin's Finches (‘Geospizines’), Orr (1945) concluded that sometimes territorial song may be used to address allospecific competitors.
Data from tyrant flycatchers further indicate that, in sympatry, allospecies songs are stored for discrimination. As for other sub-Oscines, songs of the Alder Flycatcher Empidonax alnorum and its sibling species the Willow Flycatcher E. trailli develop normally when birds are raised in isolation, indicating that imitative learning plays no part in song ontogeny in these species (Kroodsma 1984). Nevertheless, playback studies in areas of sympatry between the two flycatchers revealed that each species would respond to songs of both forms. In areas of allopatry each species would respond only to conspecific song in playback trials (Prescott 1987). The related Least Flycatcher E. minimus will store songs of American Redstarts Setophaga ruticilla in areas of sympatry and will respond aggressively to playback of allospecific songs of the latter (Martin et al. 1996).
STORAGE FOR DISCRIMINATION AND PERFORMANCE
Individual recognition
A body of literature indicates that both Oscines and non-Oscines can distinguish neighbours from strangers or different neighbours individually (Falls 1982, Stoddard 1996). Manifestations of individual recognition include dialect formation and matched countersinging by males.
Neighbour recognition
Chaffinches Fringilla coelebs are one of the best studied dialectal avian species (e.g. Baker & Jenkins 1987) and were the subject of the classical pioneering studies in song learning and vocal production (Thorpe 1958). Hinde (1958) conducted playback studies on captives using different song themes and observed that the proportion of themes matching the playback song tended to increase in song bouts following experimental trials. Field studies with Chaffinches on the shores of Lake Constance, Germany, indicated that some individuals would respond to playback themes with like themes in most trials (Fig. 2, Baptista unpublished). This phenomenon has been dubbed ‘matched countersinging’ by investigators and has been described for many Oscine species (Horn & Falls 1996).
One of the most impressive examples of matched countersinging is that found in Song Sparrows. Each male produces a song repertoire consisting of a mean of nine different themes, but consisting of at least one theme sung by each of his neighbours. Each neighbour's theme is often matched with a like theme in bouts of countersinging (Beecher 1996, Beecher et al. 1997). These data indicate that not only do some birds learn neighbours' song themes, but may discriminate between them and by matched-countersinging may ‘name’ the neighbour to which the song is directed (Armstrong 1963; Catchpole & Slater 1995).
Mate recognition
In some vocal learning species, females not only attend to the vocalisations of males but mated pairs also learn and produce accurate copies of the each other's vocalisations. Mated pairs of Cardueline Finches in a number of genera imitate contact calls (Mundinger 1970, review in Baptista & Gaunt 1997a). Males may imitate from females or vice-versa, thus enabling them to recognise each other from a distance. Female Bullfinches learn their mate's songs, enabling pairs to recognise and locate each other (Nicolai 1959).
Group or population recognition
In territorial species, songs are often used to repulse conspecifics in competitive situations. However, vocalisations may also function in attraction, e.g. calls may serve as an acoustical label of membership in a flock and aid in co-ordinating and synchronising movements of the group (Farabaugh & Dooling 1996). Experiments indicate that birds use calls or songs to identify social groups or populations to which they belong.
For example, flocks of Black-capped Chickadees Parus articapillus can be distinguished on the basis of their ‘chick-a-dee’ call. Birds from different origins housed as three captive, artificial flocks converged on each other's calls after a month together, indicating that certain parameters are learned (Mammen & Nowicki 1981). Playback studies indicated that individuals distinguished flock members from strangers based on this call (Nowicki 1983).
Similarly, flocks of Yellow-napped Amazon Parrots Amazona ochrocephala occupy exclusive roost areas that may be distinguished by their calls (Wright 1996). Budgerigars Melopsittacus undulatus also identity flock members by call with learning of the flock specific calls continuing into adulthood (Farabaugh & Dooling 1996).
Vocal acquisition through learning coupled with geographical isolation often leads to the formation of regional song dialects. Playback studies with various dialectal species, e.g. White-crowned Sparrows Zonotricia leucophrys and Corn Buntings Emberiza calandra, indicate that birds may distinguish local from alien dialects based on song (McGregor 1983, Tomback et al. 1983, Baptista & Gaunt 1997b).
Allospecies recognition
Acquisition and production of allospecific vocalisations have been described in a plethora of avian species (e.g. review in Dobkin 1979). However, use of alien sounds in communication has seldom been documented. Pink Cockatoos Cacatua leadbeateri may usurp the nests of Galahs Cacatua roseicapilla after the latter have already laid eggs. Alien and conspecific eggs are incubated and chicks are raised by the usurping parents, and, although Galahs do not imitate alarm calls from Pink Cockatoos, they do imitate and use Pink Cockatoo contact calls to maintain contact with their adopted flock members (Rowley & Chapman 1986).
The Thick-billed Euphonia Euphonia laniirostris has been documented imitating calls of the most common Passerines in their habitat in Panama and Colombia (Morton 1976, Remsen 1976). Morton (1976) noted that the Euphonia produced alarm calls of the mimicked species in the correct context and suggested that this was a device to evoke mobbing by the latter.
Mimetic species such as menurids, mimids, sturnids and ptilonorhynchids often imitate a variety of birds in their acoustical umwelt (Robinson 1975, Loffredo & Borgia 1986, Hausberger et al. 1991). However, these mimicked sounds are not used in interspecific communication. The Cockatoos and Euphonias discussed above are special cases as they actually use mimicked sounds to address allospecies.
DISCRIMINATION AND POSSIBLE PERFORMANCE
Individual recognition
Increasing evidence has accumulated demonstrating that some birds store more vocal memories than they perform and/or perform vocal memory stored early in life at a much later time. Indeed, this phenomenon makes it difficult to know with certainty if a new song in an adult bird's repertoire is newly acquired or a retrieved song from memory (Nelson et al. 1996).
Great Tits Parus major were thought to learn songs for performance during an early sensitive phase, and experiments indicated that they continued to recognise new neighbours throughout life. These data indicated that storing for discrimination is not age restricted (McGregor & Avery 1986). However, Great Tits have been shown to change their song from year to year (McGregor & Krebs 1989). This behaviour could be interpreted as retrieval of stored information or learning anew, the two phenomena being difficult to separate under natural condition.
A White-crowned Sparrow was recorded singing two dialects in May 1993, the local dialect of San Francisco Z. l. nuttalli and a dialect typical of visiting Z. l. gambelii from Alaska. In May of the following year he was recorded singing only the local dialect. To test if the unused dialect could be evoked, playback studies were performed using the Alaskan song. The bird responded to 78 playback trials with 120 local songs and four Alaskan songs (Baptista et al. 1997). This finding suggested that unused songs could be evoked if the stimulus were intense enough, e.g. aggravated attacks by an intruder singing the rare song type.
Female White-rumped Munias Lonchura striata can distinguish their father's song from those of other males in playback trials (Dietrich 1980). However, unlike their relative the Zebra Finch, in which females do not sing, a few White-rumped females have been noted singing on rare occasions (Restall 1996; Baptista, pers. obs.).
Population recognition
Female White-crowned Sparrows implanted with estradiol will perform more solicitation displays to playback of familiar local dialects than unfamiliar ones (Baker et al. 1982, Chilton & Lein 1996). Female song in this species was thought to be unusual (Konishi 1965, Baptista 1974). However, recent studies indicate that some females often sing in the non-breeding season (Baptista et al. 1993). One widowed female sang copiously, apparently advertising for a mate. A male was mated to two females who often sang at each other (Blanchard 1936). This finding indicates that although females normally do not sing, they can under the appropriate circumstances.
Allospecies recognition
A growing body of literature indicates that two competing Oscine species may store each other's songs in areas of sympatry and use these songs in discrimination, i.e. identifying the competing allospecies. Examples include both related and unrelated taxa. In Sympatric related species such as New World buntings Passerina spp., Old World Acrocephalus warblers, nightingale species Luscinia spp., playback studies revealed that they would respond aggressively to each other's songs (Emlen et al. 1975, Catchpole 1978, Sorjonen 1986).
This phenomenon has been described even for unrelated taxa, e.g. Bewick's Wrens Thryomanes bewickii and Song Sparrows (Gorton 1977), Great Tits and Chaffinches (Reed 1982), White-crowned Sparrows and Song Sparrows (Catchpole & Baptista 1988, Baptista & Catchpole 1989). Some Lazuli Buntings Passerina amoena and some Indigo Buntings P. cyanea will sing each other's songs (Emlen et al. 1975). Similarly, some Song Sparrows may sing White-crowned Sparrow songs and vice-versa (review in Baptista 1990). Irrespective of whether these buntings or emberizid Finches in sympatry sing allospecies songs, they will nonetheless respond to playback of allospecies tapes. These data indicate that they store each other's songs in sympatry, and under special circumstances will produce those alien songs.
DISCUSSION
In this review we examined avian vocal communication between adults and young or adults and adults for evidence of cognitive processes, notably those of categorisation, retrieval and discrimination. We present evidence indicating that acoustic signals are stored, recovered and used to distinguish individuals, populations or species in their umwelt (Table 1). We also presented evidence that a continuum exists between storing for discrimination only and storing for discrimination plus performance. Here we place the cognitive processes that appears to be used for vocal memory into an evolutionary, adaptive significance context.
Mutual recognition between parents and young is important in colonial (e.g. various seabirds) or semi-colonial species (e.g. Zebra Finches and Mourning Doves). Selection would be against parents provisioning young not their own as this behaviour would be inimical to individual fitness. Conversely, young seeking food from strange adults are liable to be attacked and injured.
The age at which parents and young learn to recognise each other's voices differs between species with nidifugous versus nidicolous young. Prenatal learning of voice may be selected for in species with nidifugous-altricial young, e.g. murres, because young approach returning parents who signal their arrival at the colony with calls. Prenatal learning of calls in nidifugous-semialtricial Anatids is selected for as there is a premium placed on young leaving their nest-site as a group when called by their mother. As semi-altricials, young are dependent on mothers for warmth, protection, and guidance to good food sources. Fledglings of nidicolous species may disperse and hide or mix with other young in creches, or broods may be divided between the parents: thus the importance of parent-young recognition of each. Prior to fledging, however, parents come to a fixed nest site to provision their young; in contrast to young of nidifugous species, learning of parents' voices may thus be delayed for some days after hatching, because locating young may be become difficult only after fledging.
Breeding success in monogamous species necessitates close co-operation between mated pairs (see above). Petrels tend to be monogamous and long-lived and extra-pair copulations are unknown in this group. There appears to be selection for high mate fidelity in these birds, and associated with this behaviour is selection for mate recognition by voice (Bretagnolle 1996). Mated pairs of various fruit dove species (Ptilinopus sp.) have been observed duetting either in captivity or in the wild (Baptista & Gaunt 1997b). Recognition of individual males in the wild has been documented for one dove, namely the Mourning Doves (Hitchcock et al. 1989). Recognition of mates is likely in Fruit Doves and should prove a fertile area of research in the future.
A body of literature indicates that members of flocking species have a higher chance of survival than solitary individuals (e.g. Parsons & Baptista 1980). Flocks often exclude strangers from the group and may occupy exclusive home ranges (e.g. Mewaldt 1964). Group specific calls that are inherited (quail) or learned (Amazon parrots, chickadees) would thus enable members to recognise and locate each other and could be selected for.
The above discussion treats groups that store acoustical information used only in discrimination, or species that discriminate and perform. Those that learn for discrimination only include non-Oscines and sub-Oscines (Tyrranidae) as well as females of some Oscine species. Adults of both sexes among various non-Oscines as well as Oscines learn to recognise their young by sound, although adults do not produce those sounds.
Learning for discrimination and performance has been documented for two of the three groups known to acquire vocalised sounds by imitative learning, namely the Oscines and parrots. Recently, matched-countersinging has been documented for two hummingbird species (Fig. 3 unpublished, and Gaunt et. al 1994). Individual recognition should thus be expected in hummingbirds (Trochilidae), the third group known to imitate sounds.
Matched-countersinging implies the ability to categorise (Horn & Falls 1996), as it requires individuals to distinguish between various themes (between-class discrimination) as well as variation within themes (within-class discrimination) that they select from their own repertoire. This task may be as simple as selecting from two themes as in White-crowned Sparrows (Chilton & Lein 1996, Baptista et al. 1997) and chaffinches (this paper), or up to over 100 themes as in Marsh Wrens Cistothorus palustris in their leader-follower song matching bouts (Kroodsma 1979).
Matched-countersinging is not the only evidence for the cognitive processes involved in (individual) neighbour recognition, however, as playback studies have shown that various birds may recognise their neighbours' songs irrespective of whether or not they perform those songs (McGregor & Avery 1986, Stoddard 1996). Neighbours may be recognised in species with single song repertoires such as the Common Yellowthroat (Geothlypis trichas, Wunderle 1978) or with repertoires of up to 175 as in the European Robin (Erithacus rubecula, Hoezel 1986, review in Stoddard 1996).
The complexity of the task of categorisation and discrimination varies greatly among species, depending on vocal repertoire, context, or sex. Petrel males respond to playback of male calls and females to female calls (Bretagnolle 1996). Thus they must discriminate between only two classes of calls in sexual recognition but must categorize a number of calls in individual recognition. Neighbour-neighbour distinction is a more complex task than neighbour-stranger recognition as it may imply within class discrimination of themes that are very similar between males, thus presupposing the ability to discern minute differences between song renditions.
Various Oscines may store more song themes in memory than they vocalise as adults; however, some of these themes may be uttered during the practice or rehearsed song stage, a phenomenon termed ‘overproduction’ (Nelson & Marler 1994, Nelson et al. 1996). The immediate neighbours with whom a young bird countersings with determines which songs are ultimately used in the vocal repertoire (Baptista & Morton 1988). However, prolonged playback, simulating an intense encounter, may sometimes evoke the performance of an unused (stored) song (e.g., Baptista et al. 1997). This finding indicates that, although investigators have repeatedly reported that Oscines store songs for discrimination only, the possibility exists that stored songs may be vocalised on rare occasions such as when encounters are intense.
Study of female song behaviour is at its infancy because females tend to be neglected as subjects of investigation (review in Baptista & Gaunt 1994). This situation probably reflects a temperate-latitude bias by investigators as most females of temperate avian species seldom or never sing, whereas many tropical females sing and duet (Levin 1996). As described in the section on discrimination without vocal performance, female Zebra Finch, Brown-headed Cowbirds and White-crowned Sparrows do give social signals to preferred populations and individuals. In addition, females of the sedentary White-crowned Sparrow subspecies (Z. l. nuttalli) may sing during the non-breeding season, and some individuals with repertoires have been observed to engage in matched countersing (Baptista et al. 1993), indicating that they can distinguish between neighbour's themes.
Subtle differences in cognitive abilities may exist between the sexes. For example, whereas both sexes of Adelie Penguins Pygoscelis adeliae could distinguish playback of mate's calls from controls, only males discriminated between neighbours and strangers (Speirs & Davis 1991). It has been suggested that females of some species such as White-crowned Sparrows or White-throated Sparrows Zonotrichia albicollis may be more discriminating than males (Ratcliffe & Otter 1996), because males use the same theme in territoriality and advertisement and females must thus learn to interpret the context from what is apparently the same sound signal. However, it is known that song rate in males of both these species is higher in unmated or widowed males (Blanchard 1941, Wassermann 1997). Perhaps females focus on song rate (cadence) as the cue to contextual information. Female Village Indigo Birds Vidua chalybeata choose sires based on song rate (Payne & Payne 1977); thus a precedent exists for song rate to serve as cues for female discriminative response. What little data we have indicate that females are capable of the same discriminative tasks as males. Nonetheless, females do not necessarily categorise or process these stored memories for the same end. This idea has best been demonstrated with female Brown-headed Cowbirds, who use their learned song to instruct young males, by way of non-song signals, to learn these preferred songs (King & West 1988). Clearly there are many open question worthy of detailed investigation in the area of avian cognition.
In point of fact, much of the evidence for avian cognition discussed here are from studies addressing problems unrelated to cognition and thus they do not attempt specifically to test any hypothesis related to cognitive processes. Our post facto discussion will, we hope, lend itself to refocusing attention on these studies from a cognitive perspective and develop leads to new approaches that will more directly consider song performance in birds from this perspective.
REFERENCES
Armstrong, E.A. 1963. A study of bird song. London; Oxford University Press: 248pp.
Baker, A.J. & Jenkins, P.F. 1987. Founder effect and cultural evolution of songs in an isolated population of Chaffinches, Fringilla coelebs, in the Chatham Islands. Animal. Behaviour. 35: 1793-1803.
Baker, J.A. & Bailey, E.D. 1987a. Ontogeny of the separation call in Northern Bobwhite (Colinus virginianus). Canadian Journal of Zoology 65: 1016-1020.
Baker, J.A. & Bailey, E.D. 1987b. Sources of phenotypic variation in the separation call in Northern Bobwhite (Colinus virginianus). Canadian Journal of Zoology 65: 1010-1015.
Baker, M.C., Spitler-Nabors, K.J., & Bradley, D.C. 1982. The response of female mountain White-crowned Sparrows to songs from their natal dialect and an alien dialect. Behavior Ecology and Sociobiology 10: 175-179.
Balda, R.P. & Balda, J.H. 1978. The care of young Piñon Jays (Gymnorhinus cyanocephalus) and their integration into the flock. Journal für Ornithologie 119: 146-171.
Baptista, L.F. 1974. The effects of songs of wintering White-crowned Sparrows on song development in sedentary populations of the species. Zietschrift für Tierpsychologie. 34: 147-171.
Baptista, L.F. 1990. Song learning in the White-crowned Sparrow (Zonotrichia leucophrys): sensitive phases and stimulus filtering revisited. Proceedings of the International 100th Deutsche Ornithological Gesellshaft Meeting. Bonn; Current Topics in Avian Biology: 143-152.
Baptista, L.F. 1996. Nature and its nurturing in avian vocal development. In: Kroodsma, D.E. & Miller, E. H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY; Cornell University Press: 39-60.
Baptista, L.F., Bell, D.A., & Trail, P.W. 1993. Song learning and production in the White-crowned Sparrow: parallels with sexual imprinting. In: ten Cate, C., Slater, P.J.B., & Kruijt, P.J. (eds) Netherlands Journal of Zoology, Proceedings International Conference on Song Learning and Imprinting: An Inquiry into Mechanisms of Behaviourial Development. 43: 17-33.
Baptista, L.F. & Catchpole, C.K. 1989. Vocal mimicry and interspecific aggression in songbirds: experiments using White-crowned Sparrow imitation of Song Sparrow song. Behaviour 109: 247-257.
Baptista, L.F. & Gaunt, S.L.L. 1994. Advances in studies of avian sound communication. Condor 96: 817-830.
Baptista, L.F. & Gaunt, S.L.L. 1997a. Social interaction and vocal development in birds. In: Hausberger, M. & Snowdon, C. (eds) Social influences on vocal development. Cambridge; Cambridge University Press: 23-40.
Baptista, L.F. & Gaunt, S.L.L. 1997b. Bioacoustics as a tool in conservation studies. In: Clemmons, J.R. & Buchholz, R. (eds) Behavioral approaches to conservation in the wild. Cambridge; Cambridge University Press: 212-242.
Baptista, L.F., Jesse, A., Bell, D., & Cebrian, C. 1997. Acquisition and recall of Gambel's Sparrow dialects by Nuttall's White-crowned Sparrows in the wild. Wilson Bulletin 109: 516-521.
Baptista, L.F. & Morton, M.L. 1988. Song learning in montane White-crowned Sparrows: from whom and when. Animal Behaviour 36: 1753-1764.
Baptista, L.F., Trail, P.W., DeWolfe. B.B., & Morton, M.L. 1993. Singing and its functions in female White-crowned Sparrows. Animal Behaviour 46:511-524.
Beecher, M.D. 1996. Birdsong learning in the laboratory and field. In: Kroodsma, D.E. & Miller E.H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY; Cornell University Press: 61-78.
Beecher, M.D., Beecher, I.M., & Hahn, S. 1981. Parent-offspring recognition in Bank Swallows (Riparia riparia): II development and acoustic basis. Animal Behaviour 29: 95-101.
Beecher, M.D., Stoddard, P.K., Campbell, S.E., & Horning, C. L. 1997. Repertoire matching between neighbouring Song Sparrows. Animal Behaviour 51: 917-923.
Blanchard, B. 1936. Continuity of behavior in the Nuttall White-crowned Sparrow. Condor 38: 145-150.
Blanchard, B. 1941. The White-crowned Sparrows (Zonotrichia leucophrys) of the Pacific seaboard: environment and annual cycle. University of California Publications in Zoology 46: 1-178.
Blockstein, D.E. & Westmoreland, D.A. 1993. Reproductive strategy. In: Baskett et al. (ed) Ecology and management of the Mourning Dove. Harrisburg, Pa.; Stackpole Books: 105-116.
Bretagnolle, V. 1996. Acoustic communication in a group of nonpasserine birds, the petrels. In: Kroodsma, D.E. & Miller, E. H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY.; Cornell University Press: 160-177.
Catchpole, C.K. 1978. Interspecific territorialism and competition in Acrocephalus Warblers as revealed by playback experiments in areas of sympatry and allopatry. Animal Behaviour 26: 1072-1080.
Catchpole, C.K. & L.F. Baptista, L.F. 1988. A test of the competition hypothesis of vocal mimicry, using Song Sparrow imitation of White-crowned Sparrow song. Behaviour 106: 119-128.
Catchpole, C.K. & Slater, P.J.B. 1995. Bird song: biological themes and variations. Cambridge; Cambridge University Press: 248pp.
Chilton, G. & Lein, M.R. 1996. Songs and sexual responses of female White-crowned Sparrows (Zonotrichia leucophrys) from a mixed dialect population. Behaviour 133: 173-198.
Clayton, N.S. 1988. Song discrimination learning in Zebra Finches. Animal Behaviour 36: 1016-1024.
Clayton, N.S. 1990a. Mate choice and pair formation in Timor and Australian mainland Zebra Finches. Animal Behaviour 39: 474-480.
Clayton, N.S. 1990b. The effects of cross-fostering on assortative mating between Zebra Finch subspecies. Animal Behaviour 40: 1102-1110.
Dietrich, K. 1980. Vorbildwahl in der Gesangsentwicklung beim Japanischen Mövchen (Lonchura striata var. domestica, Estrildidae). Zietshrift für Tierpsychologie 52: 57-76.
Dobkin, D.S. 1979. Functional and evolutionary relationships of vocal copying phenomena in birds. Zeitschrift für Tierpsychologie 50: 348-363.
Emlen, S.T., Rising, J.D., & Thompson, W.L. 1975. A behavioral and morphological study of sympatry in the Indigo and Lazuli Buntings of the Great Plains. Wilson Bulletin 87: 145-179.
Falls, J.B. 1982. Individual recognition by sound in birds, In: Kroodsma, D.E. & Miller, E.H. (eds) Acoustic communication in birds 2: 237-278. New York; Academic Press.
Farabaugh, S.M. & Dooling, R.J. 1996. Acoustic communication in parrots: laboratory and field studies of Budgerigars, Melopsittacus undulatus, In: D.E. & Miller, E.H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY; Cornell University Press: 97-117
Gaunt, S.L.L., Baptista, L.F., Sánchez, J.E. & Hernandez, D. 1994. Song learning as evidenced from song sharing in two Hummingbird species (Colibri coruscans and C. thalassinus. Auk 111: 87-103.
Goldstein, R.B. 1978. Geographic variation in the ‘hoy’ call of the Bobwhite. Auk 95: 85-94.
Gorton Jr., R.E. 1977. Territorial interactions in sympatric Song Sparrows and Bewick's Wren populations. Auk 94: 701-708.
Hamilton, K.S., King, A.P., Sengelaub, D.R. & West, M.J. 1997. A brain of her own: A neural correlate of song assessment in a female songbird. Neurobiology Learning and Memory 68: 325-332.
Hausberger, M., Jenkins, P.F., & Keene, J. 1991. Species-specificity and mimicry in bird song: are they paradoxes? A reevaluation of song mimicry in the European Starling. Behaviour 117: 5 3-81.
Hinde, R.A. 1958. Alternative motor patterns in Chaffinch song. Animal Behaviour 6: 211-218.
Hitchcock, R.R., Mirarchi, R.E., & Lishak, R.S. 1989. Recognition of individual male parent vocalizations by nestling Mourning Doves. Animal Behaviour 37: 517-519.
Hoezel, A.R. 1986. Song characteristics and response to playback of male and female Robins, Erithacus rubecula. Ibis 128: 115-127.
Horn, A.G. & Falls, J.B. 1996. Categorization and the design of signals: the case of song repertoires, In: D.E. & Miller, E.H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY; Cornell University Press: 121-135.
Immelmann, K. 1962. Beträge zu einer vergleichenden Biologie australische Prachtfinken (Spermestidae). Zoologische Jahrbücher Abteilung für Systematik Ökologie und Geographie der Tiere 90: 1-196.
Johnston, T.D. 1988. Developmental explanation and the ontogeny of birdsong: nature/nurture redux. Behavioral and Brain Science 11: 617-630.
Jouventin, P., Guillotin, M., & Cornet, A. 1979. Le chant du Manchot Empereur et sa signification adaptive. Behaviour 70: 231-250.
King, A.P. & West, M.J. 1988. Searching for the functional origins of cowbird song in eastern Brown-headed Cowbirds (Molothrus ater ater). Animal Behaviour 36: 1575-1588.
King, A.P. & West, M.J. 1990. Variation in species typical behavior: A contemporary theme for comparative psychology. In: Dewsbury, D.A. (ed), Contemporary issues in comparative psychology. Sunderland, MA; Sinauer: 331-339.
Kamil, A.C. 1994. A synthetic approach to the study of animal intelligence. In: Real, L.A. (ed) Behavioral mechanisms in evolutionary biology. Chicago; University of Chicago Press: 11-45.
Konishi, M. 1965. The role of auditory feedback in the control of vocalization in the White-crowned Sparrow. Zeitschrift für Tierpsychologie 22: 770-783.
Kress, S.W. 1983. The use of decoys, sound recordings & gull control for re-establishing a tern colony in Maine. Colonial Waterbirds 6:185-196.
Kress, S.W. & Nettleship, D.N. 1988. Re-establishment of Atlantic Puffins (Fratercula arctica) at a former breeding site in the Gulf of Maine. Journal of Field Ornithology 59:161-170.
Kroodsma, D.E. 1979. Vocal dueling among male Marsh Wrens: Evidence for ritualized expressions of dominance/subordinance. Auk 96:506-515.
Kroodsma, D.E. 1984. Songs of the Alder Flycatcher (Empidonax alnorum) and Willow Flycatcher (Empidonax traillii) are innate. Auk 101: 13-24.
Levin, R.N. 1996. Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus. 1. Removal experiments. Animal Behaviour 52: 1093-1106.
Loffredo, C.A. & Borgia, G. 1986. Male courtship vocalizations as cues for mate choice in the Satin Bowerbird (Ptilonorhynchus violaceus). Auk 103: 189-195.
Luther, D.M. 1979. An intensive study of parental behavior in the Mourning Dove. Indiana Audubon Quarterly 57: 209-232.
Mammen, D.L. & Nowicki, S. 1981. Individual and within-flock convergence in chickadee calls. Behavior Ecology and Sociobiology 9: 179-186.
Marler, P. 1987. Sensitive periods and the roles of specific and general sensory stimulation in birdsong learning. In: Rauschecker, J.P. & Marler, P. (eds) Imprinting and cortical plasticity. New York; John Wiley & Sons: 99-135.
Martin, P.R., Fotheringham, J.R., Ratcliffe L., & Robertson, R.J. 1996. Response of American Redstarts (suborder Passeri) and Least Flycatchers (suborder Tyranni) to heterospecific playback: the role of song in aggressive interactions and interference competition. Behavior Ecology and Sociobiology 39: 227-235.
McArthur, P.D. 1982. Mechanisms and development of parent-young recognition in the Piñon Jays (Gymnorhinus cyanocephalus). Animal Behaviour 30: 62-74.
McGregor, P.K. 1983. The response of Corn Buntings to playback of dialects. Zeitschrift für Tierpsychologie 62: 256-260.
McGregor, P.K. & Avery, M.I. 1986. The unsung songs of Great Tits (Parus major): learning neighbour's songs for discrimination. Behavior Ecology and Sociobiology 18: 311-316.
McGregor, P.K. & Krebs, J.R. 1989. Song learning in adult Great Tits (Parus major): effects of neighbours. Behaviour 108: 139-159.
Mewaldt, L.R. 1964. Effects of bird removal on a winter population of sparrows. Bird-Banding 35: 184-195.
Miller, D.B. 1979. Long-term recognition of father's song by female Zebra Finches. Nature 280: 389-391.
Miller, D.B. & Gottlieb, G. 1976. Acoustic features of Wood Duck (Aix sponsa) maternal calls. Behaviour 57: 260.
Morton, E.S. 1976. Vocal mimicry in the Thick-billed Euphonia. Wilson Bulletin 88: 485-487.
Moscovitch, M. 1995. Models of consciousness and memory. In: Gazzaniga, M.S. (ed.) The cognitive neurosciences. Cambridge, MA; MIT Press: 1341-1356.
Mundinger, P. 1970. Vocal imitation and individual recognition of finch calls. Science 168:480-482.
Nelson, D.A. & Marler, P. 1994. Selection-based learning in bird song development. Proceedings of the National Academy of Sciences (U.S.A.) 91:10498-10501.
Nelson, D.A., Marler, P. & Morton, M.L. 1996. Overproduction in song development: an evolutionary correlate with migration. Animal Behaviour 51: 1127-1140.
Nicolai, J. 1959. Familientradition in der Gesangsentwicklung des Gimpels (Pyrrhula pyrrhula L.). Journal für Ornithologie 100: 39-46.
Nowicki, S. 1983. Flock-specific recognition of Chickadee calls. Behavior Ecology and Sociobiology 12: 317-320.
Orr, R.T. 1945. A study of captive Galápagos Finches of the genus Geospiza. Condor 47: 177-201.
Parsons, J. & Baptista, L.F. 1980. Crown color and dominance in the White-crowned Sparrow. Auk 97: 807-815.
Payne, R.B. & Payne, K. 1977. Social organization and mating success in local song populations of Village Indigobirds (Vidua chalybeata). Zeitschrift für Tierpsychologie 45: 113-173.
Podolsky, R.H. 1990. Effectiveness of social stimuli in attracting Laysan albatross to new potential nesting sites. Auk 107:119-125.
Podolsky, R.H. & Kress, S.W. 1992. Attraction of the endangered Dark-rumped Petrel to recorded vocalizations in the Galapagos Islands. Condor 94: 448-453.
Prescott, D.R.C. 1987. Territorial responses to song playback in allopatric and sympatric populations of Alder (Empidonax alnorum) and Willow (E. traillii) flycatchers. Wilson Bulletin 99:611-619.
Ratcliffe, L. & Otter, K. 1996. Sex differences in song recognition. In: Kroodsma, D.E. & Miller, E.H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY; Cornell University Press: 339-355.
Reed, T.M. 1982. Interspecific territoriality in the Chaffinch and Great Tit on islands and the mainland of Scotland: playback and removal experiments. Animal Behaviour 30: 171-181.
Remsen, J.V. 1976. Observations of vocal mimicry in the Thick-Billed Euphonia. Wilson Bulletin 88: 487-488.
Restall, R. 1996. Munias and mannikins. Sussex; Pica Press: 264pp.
Robinson, F.N. 1975. Vocal mimicry and the evolution of bird song. Emu 75: 23-27.
Rowley, I. & Chapman, G. 1986. Cross-fostering, imprinting and learning in two sympatric species of cockatoo. Behaviour 96: 1-16.
Smith, J.N.M. & Merkt, J.R. 1980. Development and stability of single-parent family units in the Song Sparrow. Canadian Journal of Zoology 58:1869-1875.
Sorjonen, J. 1986. Mixed singing and interspecific territoriality -- consequences of secondary contact of two ecologically and morphologically similar nightingale species in Europe. Ornis Scandinavia 17:53-67.
Speirs, E.A. & Davis, L.S. 1991. Discrimination by Adelie Penguins Pygoscelis adeliae between the loud mutual calls of mates, neighbours and strangers. Animal Behaviour 41:937-944.
Stoddard, P.K. 1996. Vocal recognition of neighbors by territorial Passerines. In: Kroodsma, D.E. & Miller, E.H. (eds) Ecology and evolution of acoustic communication in birds. Ithaca, NY; Cornell University Press: 356-374.
Stoddard, P.K. & Beecher, M.D. 1983. Parent recognition of offspring in the Cliff Swallow. Auk 100: 795-799.
Stokes, A.W. 1967. Behavior of the Bobwhite, Colinus virginianus. Auk 84: 1-33.
Thorpe, W.H. 1958. The learning of song patterns by birds, with especial reference to the song of the Chaffinch, (Fringilla coelebs). Ibis 100: 535-570.
Tomback, D.F., Thompson, D.B. & Baker, M.C. 1983. Dialect discrimination by White-crowned Sparrows: reactions to near and distant dialects. Auk 100: 452-460.
Tschanz, B. 1968. Trottellummen. Zeitschrift für Tierpsychologie Supplement 4.
Wassermann, F.E. 1977. Mate attraction functions of song in the White-throated Sparrow. Condor 79: 125-127.
Williams, H.W. 1969. Vocal behavior of the adult California Quail. Auk 86: 631-659.
Wright, T.F. 1996. Regional dialects in the contact call of a parrot. Proceedings of the Royal Society of London (Biology) 263: 867-872.
Wunderle, J.M. 1978. Differential response of territorial Yellowthroats to the songs of neighbors and non-neighbors. Auk 95: 389-395.
Zann, R.A. 1996. The Zebra Finch. Oxford, New York; Oxford University Press: 335pp.
Table 1. Categorisation and Discrimination in various avian taxa.
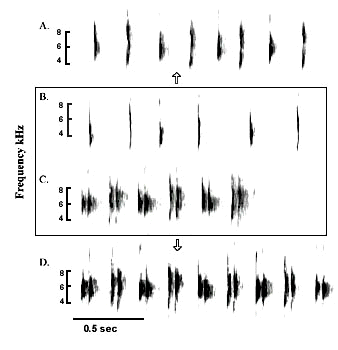
Advertising songs of three male Mourning Doves. Note the variation in frequency modulation of the last portion of the first element and the duration of all elements.
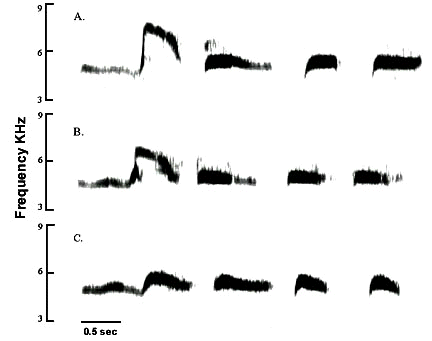
Songs of a Chaffinch from Lake Constance, Germany (A and B). The contingency table below details the number of responses to playback stimuli of the two song themes. Note the high incidences of responses with like themes (matched countersinging).
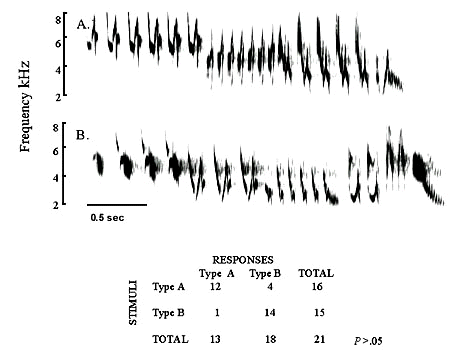
Songs of Green Violet-ears recorded at one lek in the Talamanca Mountains of Costa Rica. A and D are the first two seconds from the long songs (11+sec) of two different adults. Within the box (B and C) are two complete songs from one juvenile when singing from the song territory of each respective adult male after they vacated their perches. Note the matching of notes by all birds and of song themes with the respective adult males by the juvenile.