RT32: What were the Enantiornithes?
Evgeny N. Kurochkin1 & Cyril A. Walker2
1Paleontological Institute, 123 Profsojuznaya Street, 117868 Moscow GSP-7, Russia, fax 7 095 339 1266, e-mail enkur@paleo.ru; 2Bird Group, Natural History Museum, Tring, Herts HP23 6AP, UK
Kurochkin, E.N. & Walker, C.A. 1999. What were the Enantiornithes? In: Adams, N.J. & Slotow, R.H. (eds) Proc. 22 Int. Ornithol. Congr., Durban: 3219-3222. Johannesburg: BirdLife South Africa.
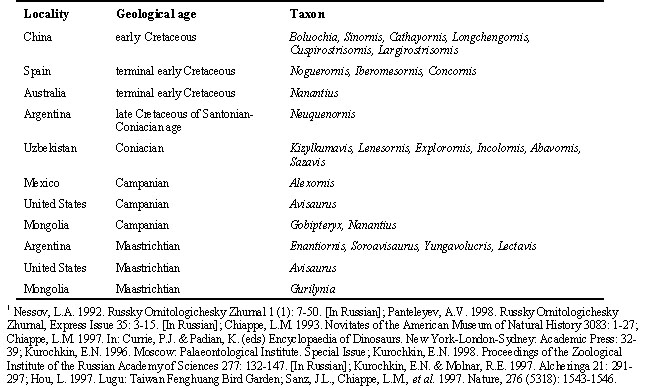
Some twenty Congress members participated in this Round Table Discussion. Unfortunately many avian Mesozoic paleontologists, who were invited to this Round Table, were not able to attend the Congress. At the beginning of this discussion E. Kurochkin presented his view on the problem of the Enantiornithes. The Enantiornithes were described some 20 years ago from the Late Cretaceous of Argentina by Walker (Walker, C.A. 1981.Nature, 292 (1): 51-53). Walker gave a very exact scientific name to them, that means ‘opposite birds’. The first diagnosis for the Enantiornithes was also correct and later confirmed by other researchers. Enantiornithes are recorded world-wide from the beginning to the end of the Cretaceous. Conceivably they might represent the most diversified and important group of Cretaceous birds. To date, about 35 forms of enantiornithine birds have been described (Table 1).
Throughout the Cretaceous, between 135 and 65 million years ago, the enantiornithines had a number of very different life forms. Small toothed arboreal enantiornithines lived in the early Cretaceous of China, Mongolia, and Spain; a small toothless arboreal form lived at the end of the early Cretaceous in Australia; small toothless coastline and arboreal forms lived in the late Cretaceous in Mongolia and Argentina; and large swimming, running and wading enantiornithines in the late Cretaceous of Argentina, North America and Mongolia. All known enantiornithines were good fliers. The early Cretaceous enantiornithines had toothed jaws, and late Cretaceous ones did not. All enantiornithines had wing digits with claws.
It has been suggested that the Enantiornithes originated in Laurasia, perhaps in the Jurassic, and that they may have restricted the wide dispersion and diversification of the Neornithes before the end of the Cretaceous in the northern Hemisphere. It is thought, that the extinction of the Enantiornithes occurred at the end of the Cretaceous, because well known Paleocene and Eocene avifaunas in Eurasia and North America lacked these birds. However, it could be possible that some relict enantiornithines could have survived through the Cretaceous/Tertiary boundary in Australia or South America.
The Enantiornithes have a very specific morphology that faultlessly differentiates them ornithurine birds. However, the taxonomic rank and relationships of Enantiornithes remain controversial. There are three competitive hypotheses: (1) Subclass Enantiornithes represents a separate lineage in avian evolution. (2) Superorder Enantiornithes represents just a primitive taxon of Ornithothoraces with respect to Ornithurae. (3) Infraclass Enantiornithes belongs to subclass Sauriurae and represents the closest relatives of Archaeopteryx.
The structural and functional morphology of the enantiornithines is a most intriguing problem. At a first superficial view, they have the same morphology of a stretched coracoid, sable-shaped scapula, pectoral-crested humerus, bowed femur, long and thin tibiotarsus, consolidated tarsometatarsus, and a long pygostyle, as do the living Ornithurae. However, a closer view shows many other details which are only characteristic of the Enantiornithes. For example, they have an articulation between a projected process (boss) on the coracoid and a concave facet (socket) on the scapula, that is completely opposite to the type of the articulation among the Ornithurae. The articulation head of the humerus (caput humeri) occupies a mid-position in the Enantiornithes, but it is displaced ventrally in the Ornithurae. In the enantiornithines the articulation in the tarsal joint lies between the proximal tarsals and the fused proximal ends of the metatarsals. Moreover, here the distal tarsals are very small or reduced, since they do not participate in the articulation. There is also a very different intertarsal articulation (between the proximal and distal tarsals) in the Ornithurae. A long pygostyle is a main feature that causes some avian experts to associate the Enantiornithes with the Ornithurae. However, this long enantiornithine pygostyle shows a probable attachment of tail feathers along it, as in Archaeopteryx, but not in the fan-shaped form present in the Ornithurae. Further similar differences might be found in the structure of the neck vertebrae, ilium, distal end of the humerus, proximal end of the ulna, shaft of the radius, distal end of the femur, and in the wing and pedal digits.
I regard the similarity between the Enantiornithes and Ornithurae as functional analogies, and not shared derived characters. It might be hypothesised that the Enantiornithes represent a side branch of avian evolution. In general, the Enantiornithes are a key group to understanding the early evolution of birds. In contrast, Chiappe et al. (Chiappe, L., Norell, M. & Clark, J. 1996. Memoirs of the Queensland Museum 39 (3): 557-582) and Chiappe (Chiappe, L.M. 1995. Nature, 378: 349-355) accepts all similar characters as homologous uniting the Enantiornithes and Ornithurae. They also claim all similar characters of Archaeopteryx, Alvarezsauridae, Enantiornithes, and Neornithes as homologous. In their cladograms the Enantiornithes occupies the basal position to the Ornithurae, whereas the last three clades are united in taxon Ornithothoraces. On such cladograms Archaeopteryx is a sister clade to the Metornithes, which contains the Ornithothoraces and maniraptoran Alvarezsauridae (Mononykus and his South American and other Mongolian relatives). Elzanowski (Elzanowski, A. 1995. Courier Forshungsinstitut Senckenberg 181: 37-53) and Chatterjee (Chatterjee, S. 1997. Baltimore and London: Johns Hopkins University Press) have similar conclusions uniting the Enantiornithes and Ornithurae. I presented evidence that similarity between the Enantiornithes and Neornithes is homoplasy in many characters. It does not support their common origin, although they developed flying abilities in parallel.
The relationships of branches of Mesozoic birds and their probable ancestors have been discussed widely with contradictory viewpoints by recent avian paleontologists. The principal point of discussion is the relationship of the Enantiornithes to the Archaeornithes, and to Archaeopteryx. Along with Larry Martin, I argue for the sister group relationship between these two lineages. There is a set of derived characters that support such a conclusion. They are the cranio-caudal depression on the cranial side of proximal humeral head, a distinct dorsal process on the ischium, elongation of the cranial portion of the ilium, additional trochanter of the proximal head of femur, caudo-lateral crest of distal head of femur, nearly complete reduction of the distal tarsals, transversal fusion of the proximal ends of the metatarsals, and the tarsal junction between proximal tarsals (fused with tibia) and a transversal fusion of proximal metatarsals. A new comparison of the skulls in Archaeopteryx and Cathayornis showed the advanced similarity in structure of the quadrate, nasal, lacrimal bones, and preorbital foramen (Martin, L.D. & Zhou, Z. 1997. Nature 389 (6551): 556). Thus, the enantiornithines and Archaeopteryx have a common origin and must be linked in the subclass Sauriurae, which is not in a sister group relationship with the Ornithurae (Hou, L., Martin, L.D., Zhou, Zhonghe & Feduccia, A. 1996. Science, 274 (5290): 1164-1167; and others). Confuciusornis well supports the unity of Archaeopteryx and Enantiornithes.
It is very possible that the difference in evolutionary pathways between the Sauriurae and Ornithurae is supported by the developmental data on the manus. Of course, the embryology of Archaeopteryx remains unknown. However, his three wing digits must represent first, second and third digits of the manus, since they contain two, three and four phalanges according to the basic pendactyl formula of the tetrapods. Reduction of the digits from the outer side is a synapomorphy of the Theropods (Hecht and Hecht, 1994). As a rule, in other tetrapods the outer fifth and inner first digits begin the reduction of the digital formula. The same way is characteristic for living birds, as it has been shown by analysis in the early developmental process (Burke, A.C. & Feduccia, A. 1997. Science, 278. (5338): 666; and others). Thus, the remaining digits of the Ornithurae are homologous with the second, third and fourth digits of the pendactyl formula. Therefore, the wing digits of Archaeopteryx and living birds are not homologous, although other points of view exist on this subject (for example see Padian, K. & Chiappe, L.M. 1998. Biol. Rev. Cambridge Phil. Soc. 73 (1): 1-42). After that, the Sauriurae represent a separate and dead lineage in the avian evolution.
The relationships of Archaeopteryx and Theropods were well supported by a number of investigations. In all likelihood, Archaeopteryx and Enantiornithes are the direct descendants of the Theropods, and a separate lineage of feathered reptiles not connected with the evolutionary lineage of the Ornithurae. Thus, birds evolved at least by two parallel independent lineages: Sauriurae and Ornithurae that probably had different ancestors. Up to recently, class Aves was considered the only vertebrate group that evolved monophyletically from some reptiles via Archaeopteryx. However, a lot of records have appeared during the last few years which indicate an unbelievable diversity of Mesozoic birds and provide a complicated understanding of early avian evolution. These data confirm that the basal evolution of birds belongs to a period long before Archaeopteryx. All main lineages of the Ornithurae have existed since the early Cretaceous. The highest avian taxa, such as the Enantiornithes, Hesperornithes, Ichthyornithes, Palaeognathae, and Neognathae existed simultaneously on the Earth throughout more than half (80 my) of the known history of birds.
After that, R. Prum, W. Boles, W. Bock, J. van den Berge, and T.Stidham delivered the speeches with some questions and opinions. They mentioned that although the unique character of the Enantiornithes and their relationship with Archaeopteryx was well evident in the presentation, a dual origin of feathers and flight from widely differing ancestors is more controversial.
Table 1: Locality, geological age and taxa of enantiornithine birds described to Science.1